
Роль устьиц листа заключается в следующем. Устьица, их строение и механизм действия. Водный обмен. лист как орган транспирации

Лист — чрезвычайно важный орган растения. Лист — часть побега. Основными функциями его являются фотосинтез и транспирация. Лист характеризуется высокой морфологической пластичностью, разнообразием форм и большими приспособительными возможностями. Основание листа может расширяться в виде косых листовидных образований — прилистников с каждой стороны листа. В некоторых случаях они настолько велики, что играют определённую роль в фотосинтезе. Прилистники бываю свободными или приросшими к черешку, они могут смещаться на внутреннюю сторону листа и тогда их называют пазушными. Основания листьев могут быть превращены во влагалище, окружающее стебель и препятствующие его сгибанию.
Внешнее строение листа
Листовые пластинки различаются по размерам: от нескольких миллиметров до 10-15 метров и даже 20 (у пальм). Продолжительность жизни листьев не превышает нескольких месяцев, у некоторых — от 1,5 до 15 лет. Размер и форма листьев являются наследственными признаками.
Части листа
Лист — боковой вегетативный орган, растущий от стебля, имеющий двустороннюю симметрию и зону роста при основании. Лист обычно состоит из листовой пластинки, черешка (исключением являются сидячие листья); для ряда семейств характерны прилистники. Листья бываю простые, имеющие одну листовую пластинку, и сложные — с несколькими листовыми пластинками (листочками).
Листовая пластинка — расширенная, обычно плоская часть листа, выполняющая функции фотосинтеза, газообмена, транспирации и у некоторых видов — вегетативного размножения.
Основание листа (листовая подушка) — часть листа, соединяющая его со стеблем. Здесь находится образовательная ткань, дающая рост листовой пластинке и черешку.
Прилистники — парные листовидные образования в основании листа. Они могут опадать при развёртывании листа или сохраняться. Защищают пазушные боковые почки и вставочную образовательную ткань листа.
Черешок — суженная часть листа, соединяющая своим основанием листовую пластинку со стеблем. Он выполняет важнейшие функции: ориентирует лист по отношению к свету, является местом расположения вставочной образовательной ткани, за счёт которой растёт лист. Кроме этого, он имеет механическое значение для ослабления ударов по листовой пластинке от дождя, града, ветра и пр.
Простые и сложные листья
Лист может иметь одну (простой), несколько или множество листовых пластинок. Если последние снабжены сочленениями, то такой лист называется сложным. Благодаря сочленениям на общем черешке листа листочки сложных листьев опадают поодиночке. Однако у некоторых растений сложные листья могут опадать и целиком.
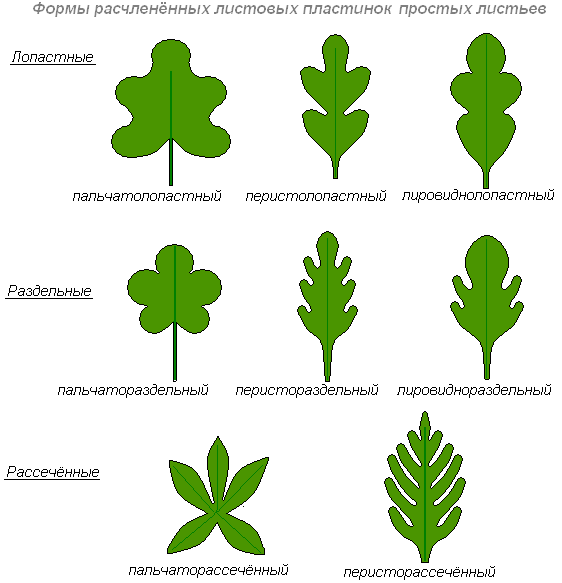
По форме цельные листья, различают как лопастные, раздельные и рассечённые.
Лопастным называю лист, у которого вырезы по краям пластинки доходят до одной четверти его ширины, а при большем углублении, если вырезы достигают более четверти ширины пластинки, лист называется раздельным. Лопасти раздельного листа называют долями.
Рассечённым называют лист, у которого вырезы по краям пластинки доходят почти до средней жилки, образуя сегменты пластинки. Раздельные и рассечённые листья могут быть пальчатые и перистые, дважды пальчатые и дважды перистые и т.д. соответственно этому различают пальчато-раздельный лист, перисторассечённый лист; непарно-перисторассечённый лист у картофеля. Он состоит из конечной доли, нескольких пар боковых долек, между которыми располагаются ещё меньшие дольки.
Если пластинка удлинённая, а доли или сегменты её треугольные, лист называют струговидным (одуванчик); если боковые доли неравновеликие, к основанию уменьшаются, а конечная доля крупная и округлая, получается лировидный лист (редька).

Что касается сложных листьев, то среди них различают тройчатосложные, пальчатосложные и перистосложные листья. Если сложный лист состоит из трёх листочков, он называется тройчатосложным, или тройчатым (клён). Если черешочки листочков прикрепляются к главному черешку как бы в одной точке, а самые листочки расходятся радиально, лист называется пальчатосложным (люпин). Если на главном черешке боковые листочки расположены с обеих сторон по длине черешка, лист называется перистосложным.
Если такой лист заканчивается наверху непарным одиночным листочком, получается, непарноперистый лист. Если же конечного нет, лист называется парноперистым.
Если каждый листочек перистосложного листа, в свою очередь, является сложным, то получается дважды перистосложный лист.
Формы цельных листовых пластинок
Сложным листом называют такой, на черешке которого имеется несколько листовых пластинок. Они крепятся к главному черешку своими собственными черешками, нередко самостоятельно, поодиночке, опадают, и называются листочками.
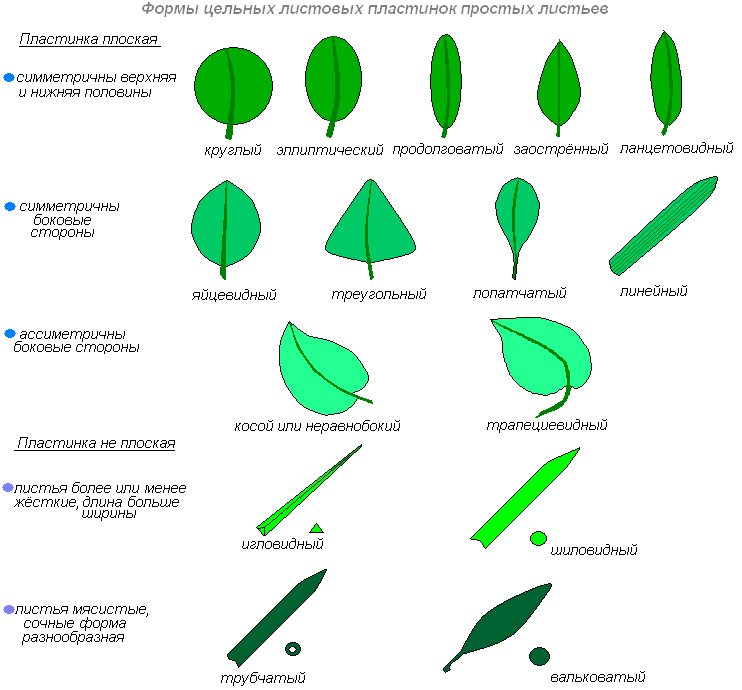
Формы листовых пластинок различных растений отличаются по очертанию, степени расчленённости, форме основания и верхушки. Очертания могут быть овальными, круглыми, эллиптическими, треугольными и другими. Листовая пластинка бывает удлиненной. Свободный конец её может быть острым, тупым, заострённым, остроконечным. Основание её сужено и оттянуто к стеблю, может быть округлым, сердцевидным.
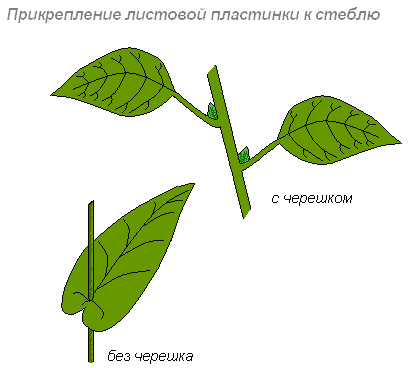
Прикрепление листьев к стеблю
Листья прикрепляются к побегу длинными, короткими черешками или бывают сидячими.
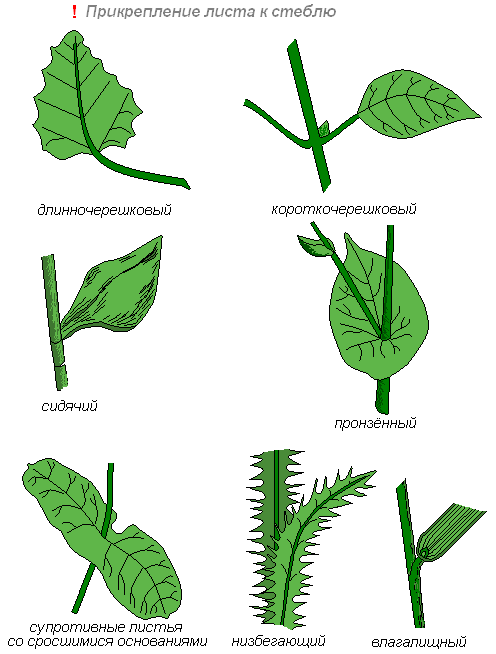
У некоторых растений основание сидячего листа на большом протяжении срастается с побегом (низбегающий лист) или побег пронизывает листовую пластинку насквозь (пронзённый лист).
Форма края листовой пластинки
Листовые пластинки различают по степени рассечённости: неглубокие надрезы — зубчатые или пальчатые края листа, глубокие вырезы — лопастные, раздельные и рассечённые края.
Если края листовой пластинки не имеют никаких выемок, лист называется цельнокрайним . Если выемки по краю листа неглубокие, лист называется цельным .
Лопастной лист — лист, пластинка которого расчленена на лопасти до 1/3 ширины полулиста.
Раздельный лист — лист с пластинкой, расчленённой до ½ ширину полулиста.
Рассечённый лист — лист, пластинка которого расчленена до главной жилки или до основания листа.
Край листовой пластинки — пильчатый (острые углы).
Край листовой пластинки — городчатый (округлые выступы).
Край листовой пластинки — выемчатый (округлые выемки).
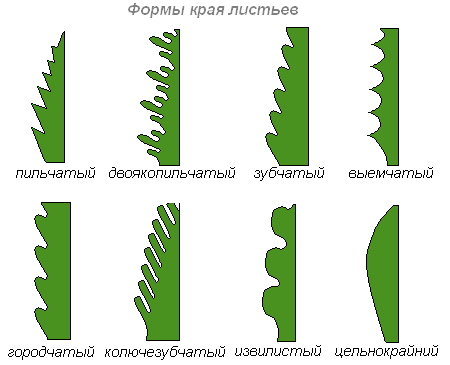
Жилкование
На каждом листе легко заметить многочисленные жилки, особенно отчётливые и рельефные на нижней стороне листа.
Жилки — это проводящие пучки, соединяющие лист со стеблем. Функции их — проводящая (снабжение листьев водой и минеральными солями и выведение из них продуктов ассимиляции) и механическая (жилки являются опорой для листовой паренхимы и защищают листья от разрывов). Среди разнообразия жилкования различают листовую пластинку с одной главной жилкой, от которой расходятся боковые ответвления по перистому или пальчатоперистому типу; с несколькими главными жилками, различающимися толщиной и направлением распределения по пластинке (дугонервный, параллельный типы). Между описанными типами жилкования существует много промежуточных или иных форм.
Исходная часть всех жилок листовой пластинки находится в черешке листа, откуда выходит у многих растений основная, главная жилка, разветвляясь потом в толще пластинки. По мере удаления от главной, боковые жилки всё утончаются. Самые тонкие большей частью находятся на периферии, а также вдали от периферии — посредине участков, окружённых мелкими жилками.
Существует несколько типов жилкования. У однодольных растений жилкование бывает дугонервным, при котором от стебля или влагалища вступает в пластинку ряд жилок, дугообразно направленных к вершине пластинки. У большинства злаков имеет место параллельнонервное жилкование. Дугонервное жилкование существует также у некоторых двудольных растений, например, подорожника. Однако и у них имеется связь между жилками.
У двудольных растений жилки образуют сильно разветвлённую сеть и соответственно этому различают жилкование сетчатонервоное, что говорит о лучшем обеспечении проводящими пучками.
Форма основания, верхушки, черешка листа
По форме верхушки пластинки листья бывают тупые, острые, заострённые и остроконечные.
По форме основания пластинки различают листья клиновидные, сердцевидные, копьевидные, стреловидные и др.
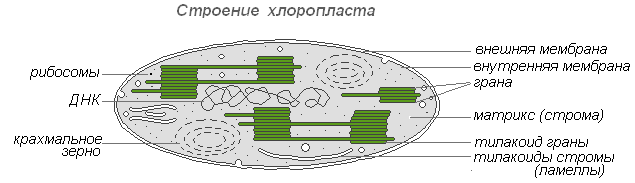
Внутреннее строение листа
Строение кожицы листа
Верхняя кожица (эпидерма) — покровная ткань на обращённой стороне листа, часто покрытая волосками, кутикулой, воском. Снаружи лист имеет кожицу (покровную ткань), которая защищает его от неблагоприятных воздействий внешней среды: от высыхания, от механических повреждений, от проникновения к внутренним тканям болезнетворных микроорганизмов. Клетки кожицы живые, по размерам и форме они разные. Одни из них более крупные, бесцветные, прозрачные и плотно прилегают друг к другу, что повышает защитные качества покровной ткани. Прозрачность клеток позволяет проникать солнечному свету внутрь листа.
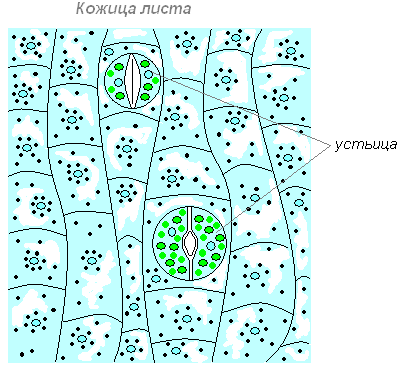
Другие клетки более мелкие, в них имеются хлоропласты, придающие им зелёный цвет. Эти клетки располагаются парами и обладают способностью изменять свою форму. При этом клетки или отдаляются друг от друга, и между ними появляется щель, или приближаются друг к другу и щель исчезает. Эти клетки назвали замыкающими, а возникающую между ними щель — устьичной. Устьице открывается, когда замыкающие клетки насыщены водой. При оттоке воды из замыкающих клеток устьице закрывается.
Строение устьица
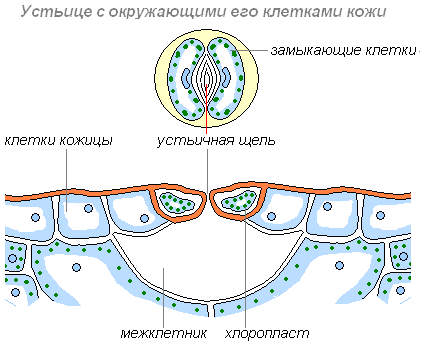
Через устьичные щели воздух поступает к внутренним клеткам листа; через них же газообразные вещества, в том числе и пары воды, выходят из листа наружу. При недостаточном обеспечение растения водой (что может случиться в сухую и жаркую погоду), устьица закрываются. Этим растения защищают себя от иссушения, так как водяные пары при закрытых устьичных щелях не выходят наружу и сохраняются в межклетниках листа. Таким образом, растения сохраняют воду в засушливый период.
Основная ткань листа
Столбчатая ткань — основная ткань, клетки которой имеют цилиндрическую форму, плотно прилегают друг к другу и расположены с верхней стороны листа (обращённой к свету). Служит для фотосинтеза. Каждая клетка этой ткани имеет тонкую оболочку, цитоплазму, ядро, хлоропласты, вакуоль. Наличие хлоропластов придаёт зелёный цвет ткани и всему листу. Клетки, которые прилегают к верхней кожице листа, вытянуты и расположены вертикально, называют — столбчатой тканью.

Губчатая ткань — основная ткань, клетки которой имеют округлую форму, расположены рыхло и между ними образуются крупные межклетники, также заполненные воздухом. В межклетниках основной ткани накапливаются пары воды, поступающие сюда из клеток. Служит для фотосинтеза, газообмена и транспирации (испарения).
Количество слоёв клеток столбчатой и губчатой тканей зависит от освещения. В листьях выросших на свету, столбчатая ткань развита сильнее, чем у листьев, выросших в условиях затемнения.

Проводящая ткань — основная ткань листа, пронизанная жилками. Жилки — это проводящие пучки, так как они образованы проводящими тканями — лубом и древесиной. По лубу осуществляется передача растворов сахара из листьев ко всем органам растения. Движение сахара идёт по ситовидным трубкам луба, которые образованы живыми клетками. Эти клетки вытянуты в длину, и в том месте, где они соприкасаются друг с другом короткими сторонами в оболочках, имеются небольшие отверстия. Через отверстия в оболочках раствор сахара переходит из одной клетки в другую. Ситовидные трубки приспособлены к передаче органического вещества на большое расстояние. Плотно по всей длине к боковой стенке ситовидной трубки прилегают живые клетки меньших размеров. Они сопутствуют клеткам трубки, и их называют клетками спутницами.
Строение жилок листа
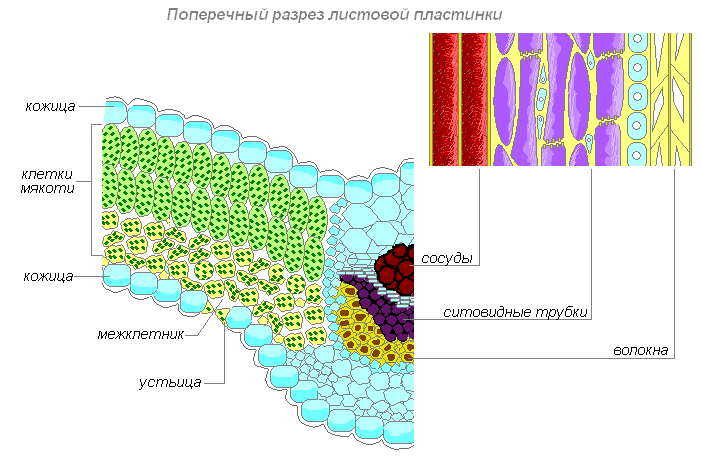
Кроме луба в состав проводящего пучка входит и древесина. По сосудам листа, так же как и в корне, движется вода с растворёнными в ней минеральными веществами. Воду и минеральные вещества растение поглощает из почвы корнями. Затем из корней по сосудам древесины эти вещества поступают в надземные органы, в том числе и к клеткам листа.
В состав многочисленных жилок входят волокна. Это длинные клетки с заострёнными концами и утолщёнными одревесневшими оболочками. Крупные жилки листа нередко окружены механической тканью, которая целиком состоит из толстостенных клеток — волокон.
Таким образом, по жилкам идёт передача раствора сахара (органического вещества) из листа к другим органам растений, а от корня — воды и минеральных веществ к листьям. Из листа растворы движутся по ситовидным трубкам, а к листу — по сосудам древесины.
Нижняя кожица покровная ткань с нижней стороны листа, обычно несёт устьица.
Жизнедеятельность листа
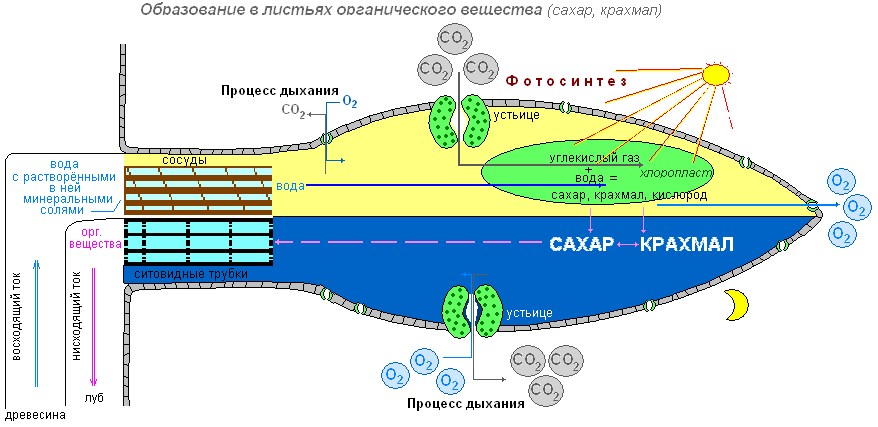
Зелёные листья — органы воздушного питания. Зелёный лист выполняет важную функцию в жизни растений — здесь образуются органические вещества. Строение листа хорошо соответствует этой функции: он имеет плоскую листовую пластинку, а в мякоти листа содержится огромное количество хлоропластов с зелёным хлорофиллом.
Вещества необходимые для образования крахмала в хлоропластах
Цель: выясним, какие вещества необходимы для образования крахмала?
Что делаем: поместим два небольших комнатных растения в тёмное место. Через два три дня первое растение поставим на кусок стекла, а рядом поместим стакан с раствором едкой щёлочи (она поглотит из воздуха весь углекислый газ), и всё это накроем стеклянным колпаком. Для того чтобы воздух не поступал к растению из окружающей среды, смажем края колпака вазелином.
Второе растение также поставим под колпак, но только рядом с растением поместим стакан с содой (или кусочком мрамора), смоченными раствором соляной кислоты. В результате взаимодействия соды (или мрамора) с кислотой выделяется углекислый газ. В воздухе под колпаком второго растения образуется много углекислого газа.
Оба растения поместим в одинаковые условия (на свет).
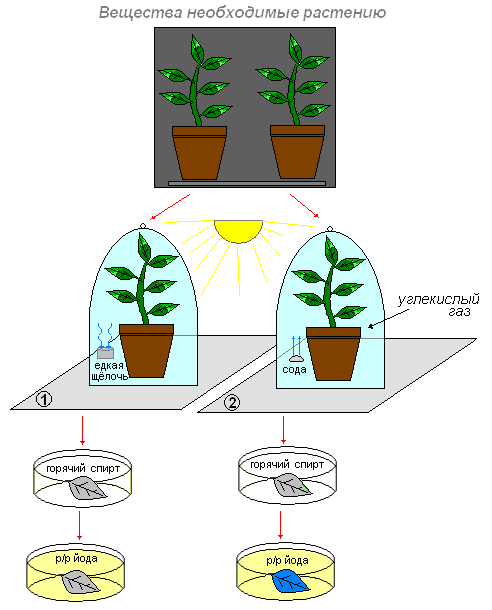
На следующий день возьмём по листу с каждого растения и обработаем вначале горячим спиртом, промываем и действуем раствором йода.
Что наблюдаем: в первом случае окраска листа не изменилась. Темно-синим стал лист того растения, которое находилось под колпаком, где был углекислый газ.
Вывод: это доказывает, что углекислый газ необходим растению для образования органического вещества (крахмал). Этот газ входит в состав атмосферного воздуха. Воздух поступает в лист через устьичные щели и заполняет пространства между клетками. Из межклетников углекислый газ проникает во все клетки.
Образование в листьях органических веществ
Цель: выяснить, в каких клетках зеленого листа образуются органические вещества (крахмал, сахар).
Что делаем: комнатное растение герань окаймлённая поместим на трое суток в тёмный шкаф (чтобы произошёл отток питательных веществ из листьев). Через трое суток вынем растение из шкафа. Прикрепим на один из листьев конверт из чёрной бумаги с вырезанным словом «свет» и поставим растение на свет или под электрическую лампочку. Через 8-10 часов срежем лист. Снимем бумагу. Опустим лист в кипящую воду, а затем на несколько минут в горячий спирт (в нём хлорофилл хорошо растворяется). Когда спирт окрасится в зелёный цвет, а лист обесцветится, промоем его водой и поместим в слабый раствор йода.
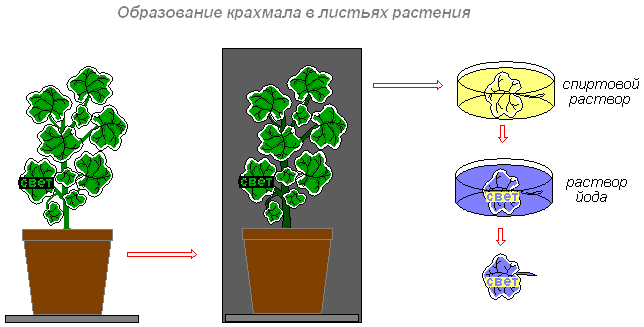
Что наблюдаем: на обесцвеченном листе появятся синие буквы (крахмал синеет от йода). Буквы появляются на той части листа, на которую падал свет. Значит, в освещённой части листа образовался крахмал. Необходимо обратить внимание на то, что белая полоска по краю листа не окрасилась. Это объясняет то, что в пластидах клеток белой полоски листа герани окаймлённой нет хлорофилла. Поэтому крахмал не обнаруживается.
Вывод: таким образом, органические вещества (крахмал, сахар) образуются только в клетках с хлоропластами, и для их образования необходим свет.
Специальные исследования учёных показали, что на свету в хлоропластах образуется сахар. Затем в результате превращений из сахара в хлоропластах образуется крахмал. Крахмал — это органическое вещество, которое в воде не растворяется.
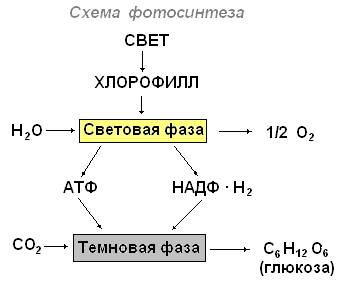
Выделяют световую и темновую фазы фотосинтеза.
Во время световой фазы фотосинтеза происходит поглощение света пигментами, образование возбуждённых (активных) молекул, обладающих избытком энергии, идут фотохимические реакции, в которых принимают участие возбуждённые молекулы пигментов. Световые реакции протекают на мембранах хлоропласта, где находится хлорофилл. Хлорофилл является высокоактивным веществом, осуществляющим поглощение света, первичное запасание энергии и дальнейшее преобразование её в химическую энергию. В фотосинтезе принимают участие и жёлтые пигменты каротиноиды.
Процесс фотосинтеза можно представить в виде суммарного уравнения:
6СО 2 + 6Н 2 О = С 6 Н 12 О 6 + 6О 2
Таким образом, суть световых реакций заключается в том, что световая энергия превращается в химическую.
Темновые реакции фотосинтеза идут в матриксе (строме) хлоропласта при участии ферментов и продуктов световых реакций и приводят к синтезу органических веществ из углекислоты и воды. Для темновых реакций не нужно непосредственное участие света.
Итогом темновых реакций является образование органических соединений.
Процесс фотосинтеза осуществляется в хлоропластах, в два этапа. В гранах (тилакоидах) протекают реакции, вызываемые светом, — световые, а в строме — реакции, не связанные со светом, — темновые, или реакции фиксации углерода.
Световые реакции
1. Свет, попадая на молекулы хлорофилла, которые находятся в мембранах тилакоидов гран, приводит их в возбуждённое состояние. В результате этого электроны ē сходят со своих орбит и переносятся с помощью переносчиков за пределы мембраны тилакоида, где и накапливаются, создавая отрицательно заряженное электрическое поле.
2. Место вышедших электронов в молекулах хлорофилла занимают электроны воды ē, так как вода под действием света подвергается фоторазложению (фотолизу):
Н 2 О↔ОН‾+Н + ; ОН‾−ē→ОН.
Гидроксилы ОН‾, став радикалами ОН, объединяются: 4ОН→2Н 2 О+О 2 , образуя воду и свободный кислород, который выделяется в атмосферу.
3. Протоны Н + не проникают через мембрану тилакоида и накапливаются внутри, используя положительно заряженное электрическое поле, что приводит к увеличению разности потенциалов по обе стороны мембраны.
4. При достижении критической разности потенциалов (200 мВ) протоны Н + устремляются по протонному каналу в ферменте АТФ-синтетаза, встроенному в мембрану тилакоида, наружу. На выходе из протонного канала создаётся высокий уровень энергии, которая идёт на синтез АТФ (АДФ+Ф→АТФ) . Образовавшиеся молекулы АТФ переходят в строму, где участвуют в реакциях фиксации углерода.
5. Протоны Н + , вышедшие на поверхность мембраны тилакоида, соединяются с электронами ē, образуя атомарный водород Н, который идёт на восстановление переносчиков НАДФ + : 2ē+2Н + =НАДФ + →НАДФ∙Н 2 (переносчик с присоединённым водородом; восстановленный переносчик).
Такими образом, активированный световой энергией электрон хлорофилла используется для присоединения водорода к переносчику. НАДФ∙Н2 переходит в строму хлоропласта, где участвует в реакциях фиксации углерода.
Реакции фиксации углерода (темновые реакции)
Осуществляется в строме хлоропласта, куда поступают АТФ, НАДФ∙Н 2 от тилакоидов гран и СО 2 из воздуха. Кроме того, там постоянно находятся пятиуглеродные соединения — пентозы С 5 , которые образуются в цикле Кальвина (цикл фиксации СО 2), Упрощённо этот цикл можно представить следующим образом:
1. К пентозе С 5 присоединяется СО 2 , в результате чего появляется нестойкое шестиугольное соединение С 6 , которое расщепляется на две трёхуглеродные группы 2С 3 — триозы.
2. Каждая из триоз 2С 3 принимает по одной фосфатной группе от двух АТФ, что обогащает молекулы энергией.
3. Каждая из триоз 2С 3 присоединяет по одному атому водорода от двух НАДФ∙Н2.
4. После чего одни триозы объединяются, образуя углеводы 2С 3 → С 6 → С 6 Н 12 О 6 (глюкоза).
5. Другие триозы объединяются, образуя пентозы 5С 3 →3С 5 , и вновь включаются в цикл фиксации СО 2 .
Суммарная реакция фотосинтеза:
6СО 2 +6Н 2 О хлорофилл энергия света →С 6 Н 12 О 6 +6О 2
Кроме углекислого газа в образовании крахмала принимает участие вода. Её растение получает из почвы. Корни поглощают воду, которая по сосудам проводящих пучков поднимается в стебель и далее в листья. А уже в клетках зелёного листа, в хлоропластах, из углекислого газа и воды при наличии света образуется органическое вещество.
Что происходит с органическими веществами, образованными в хлоропластах?
Образовавшийся в хлоропластах крахмал под воздействием особых веществ превращается в растворимый сахар, который поступает к тканям всех органов растения. В клетках некоторых тканей сахар может вновь превратиться в крахмал. Запасной крахмал накапливается в бесцветных пластидах.
Из сахаров, образовавшихся при фотосинтезе, а также минеральных солей, поглощённых корнями из почвы, растение создаёт вещества, которые ему необходимы: белки, жиры и многие другие белки, жиры и многие другие.
Часть органических веществ, синтезированных в листьях, расходуется на рост и питание растения. Другая часть откладывается в запас. У однолетних растений запасные вещества откладываются в семенах, плодах. У двулетних на первом году жизни они накапливаются в вегетативных органах. У многолетних трав вещества запасаются в подземных органах, а у деревьев и кустарников — в сердцевине, основной ткани коры и древесины. Кроме того, у них на определённом году жизни органические вещества начинают запасаться также в плодах и семенах.
Типы питания растения (минеральное, воздушное)
В живых клетках растения постоянно происходит обмен веществ и энергии. Одни вещества поглощаются и используются растением, другие выделяются в окружающую среду. Из простых веществ образуются сложные. Сложные органические вещества расщепляются на простые. Растения накапливает энергию, а в процессе фотосинтеза и освобождает её при дыхании, используя эту энергию для осуществления различных процессов жизнедеятельности.
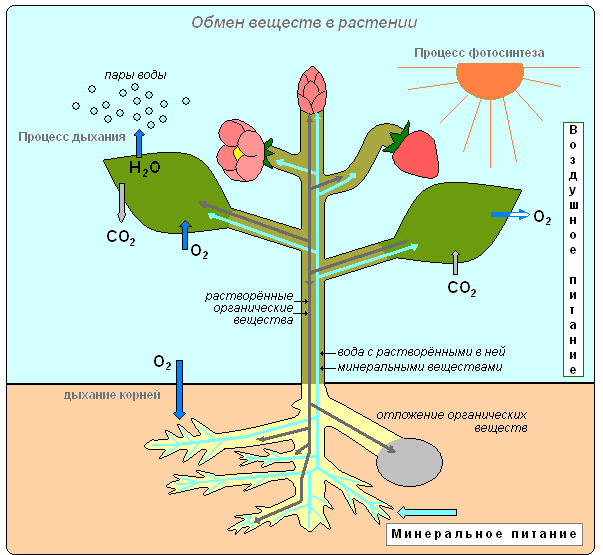
Газообмен
Листья благодаря работе устьиц осуществляют и такую важную функцию, как газообмен между растением и атмосферой. Через устьица лист с атмосферным воздухом поступают углекислый газ и кислород. Кислород используется при дыхании, углекислый газ необходим растению для образования органических веществ. Через устьица в воздух выделяется кислород, который образовался в процессе фотосинтеза. Удаляется и углекислый газ, появившийся у растения в процессе дыхания. Фотосинтез осуществляется только на свету, а дыхание на свету и в темноте, т.е. постоянно. Дыхание во всех живых клетках органов растения происходит непрерывно. Как и животные, растения погибают с прекращением дыхания.
В природе происходит обмен веществ между живым организмом и окружающей средой. Поглощение растением одних веществ из внешней среды сопровождается выделением других. Элодея, будучи водным растением, использует для питания углекислый газ, растворённый в воде.
Цель: выясним, какое же вещество выделяет элодея во внешнюю среду при фотосинтезе?
Что делаем: стебли веточек подрежем под водой (вода кипяченная) у основания и прикроем стеклянной воронкой. Пробирку, до краёв заполненную водой помещаем на трубку воронки. Это сделать в двух вариантах. Одну ёмкость поставить в тёмное место, а другую — выставить на яркий солнечный или искусственный свет.
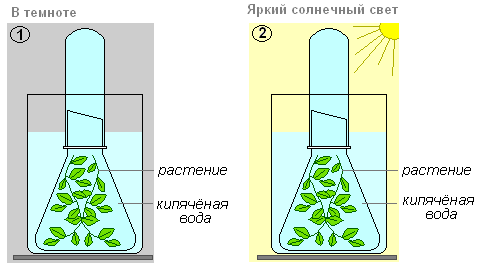
В третью и четвёртую ёмкости добавить углекислый газ (добавить небольшое количество питьевой соды или можно подышать в трубочку) и так же один поставить в темноту другой на солнечный свет.

Что наблюдаем: через некоторое время в четвёртом варианте (сосуд, стоящий на ярком солнечном свете) начинают выделяться пузырьки. Этот газ вытесняет из пробирки воду, её уровень в пробирке вытесняется.
Что делаем: когда вода будет вытеснена газом полностью, необходимо осторожно снять пробирку с воронки. Плотно закрыть отверстие большим пальцем левой руки, а правой быстро внести в пробирку тлеющую лучинку.
Что наблюдаем: лучинка загорается ярким пламенем. Посмотрев на растения, которые поместили в темноту, увидим, что пузырьки газа из элодеи не выделяются, и пробирка осталась заполненная водой. То же самое с пробирками в первом и втором варианте.
Вывод: отсюда следует, что газ, который выделила элодея — кислород. Таким образом, растение выделяет кислород только тогда, когда есть все условия для фотосинтеза — вода, углекислый газ, свет.
Испарение воды листьями (транспирация)
Процесс испарения воды листьями у растений регулируется открыванием и закрыванием устьиц. Закрывая устьица, растение защищает себя от потери воды. Открывание и закрывание устьиц находится под влиянием факторов внешней и внутренней среды, в первую очередь температуры и интенсивности солнечного света.
Листья растений содержат много воды. Она поступает по проводящей системе от корней. Внутри листа вода продвигается по стенкам клеток и по межклетникам к устьицам, через которые уходит в виде пара (испаряется). Этот процесс легко проверить, если выполнить несложное приспособление, как показано на рисунке.

Испарение воды растением называется транспирацией. Воду испаряет поверхность листа растения, особенно интенсивно — поверхность листа. Различают транспирацию кутикулярную (испарение всей поверхностью растения) и устьичную (испарение через устьица). Биологическое значение транспирации состоит в том, что она является средством передвижения воды и различных веществ по растению (присасывающее действие), способствует поступлению углекислого газа внутрь листа, углеродному питанию растений, защищает листья от перегрева.
Интенсивность испарения воды листьями зависит от:
- биологических особенностей растений;
- условий роста (растения засушливых местностей испаряют мало воды, влажных — значительно больше; теневые растения испаряют воды меньше, чем световые; много воды растения испаряют в зной, значительно меньше — в облачную погоду);
- освещения (рассеянный свет уменьшает транспирацию на 30-40%);
- содержания воды в клетках листа;
- осмотического давления клеточного сока;
- температуры почвы, воздуха и тела растения;
- влажности воздуха и скорости ветра.
Наибольшее количество воды испаряется у некоторых видов древесных пород через листовые рубцы (рубец, оставляемый опавшими листьями на стебле), которые оказываются наиболее уязвимыми местами на дереве.
Взаимосвязь процессов дыхания и фотосинтеза
Весь процесс дыхания протекает в клетках растительного организма. Он состоит из двух этапов, в ходе которых органические вещества расщепляются на углекислый газ и воду. На первом этапе при участии специальных белков (ферментов) происходит распад молекул глюкозы на более простые органические соединения и выделяется немного энергии. Этот этап дыхательного процесса происходит в цитоплазме клеток.
На втором этапе простые органические вещества, образовавшиеся на первом этапе, под действием кислорода распадаются на углекислый газ и воду. При этом высвобождается много энергии. Второй этап дыхательного процесса протекает только с участием кислорода и в специальных тельцах клетки.
Поглощённые вещества в процессе преобразований в клетках и тканях становятся веществами, из которых растение строит своё тело. Все преобразования веществ, происходящее в организме, всегда сопровождаются потреблением энергии. Зелёное растение, как автотрофный организм, поглощая световую энергию Солнца, накапливает её в органических соединениях. В процессе дыхания при расщеплении органических веществ эта энергия высвобождается и используется растением для процессов жизнедеятельности, которые происходят в клетках.
Оба процесса — фотосинтез и дыхание — идут путём последовательных многочисленных химических реакций, в которых одни вещества преобразуются в другие.
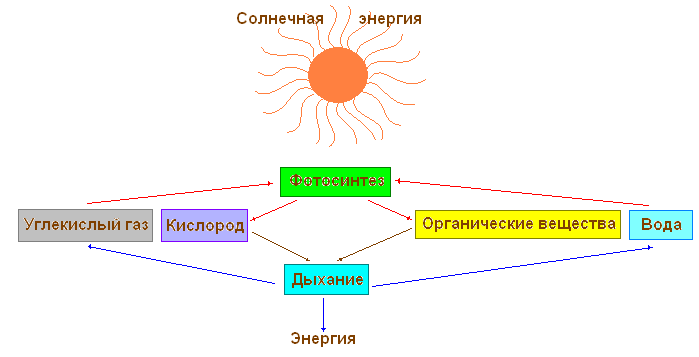
Так, в процессе фотосинтеза из углекислого газа и воды, полученных растением из окружающей среды, образуются сахара, которые затем превращаются в крахмал, клетчатку или белки, жиры и витамины — вещества, необходимые растению для питания и запасания энергии. В процессе дыхания, наоборот, происходит расщепление созданных в процессе фотосинтеза органических веществ на неорганические соединения — углекислый газ и воду. При этом растение получает высвобождающуюся энергию. Эти превращения веществ в организме называют обменом веществ. Обмен веществ — один из важнейших признаков жизни: с прекращением обмена веществ прекращается жизнь растения.
Влияние факторов среды на строение листа
Листья растений влажных мест, как правило, крупные с большим количеством устьиц. С поверхности этих листьев испаряется много влаги.
Листья растений засушливых мест невелики по размеру и имеют приспособления, уменьшающие испарение. Это густое опушение, восковой налёт, относительно небольшое число устьиц и др. У некоторых растений листья мягкие и сочные. В них запасается вода.

Листья теневыносливых растений имеют всего два-три слоя округлых, неплотно прилегающих друг к другу клеток. Крупные хлоропласты расположены в них так, что не затеняют друг друга. Теневые листья, как правило, более тонкие и имеют более тёмную зелёную окраску, так как содержат больше хлорофилла.
У растений открытых мест мякоть листа насчитывает несколько слоев, плотно прилегающих друг к другу столбчатых клеток. В них содержится меньше хлорофилла, поэтому световые листья имеют более светлую окраску. Те и другие листья иногда можно встретить и в кроне одного и того же дерева.
Защита от обезвоживания
Наружная стенка каждой клетки кожицы листа не только утолщена, но и защищена кутикулой, которая плохо пропускает воду. Защитные свойства кожицы значительно повышаются при образовании волосков, которые отражают солнечные лучи. Благодаря этому нагревание листа понижается. Всё это ограничивает возможность испарения воды с поверхности листа. При недостатке воды закрывается устьичная щель и пар не выходит наружу, накапливаясь в межклетниках, что приводит к прекращению испарения с поверхности листа. Растения жарких и сухих мест обитания имеют небольшую пластинку. Чем меньше поверхность листа, тем меньше опасность излишней потери воды.
Видоизменения листьев
В процессе приспособления к условиям окружающей среды листья у некоторых растений видоизменились потому, что стали играть роль не свойственную типичным листьям. У барбариса часть листьев видоизменились в колючки.

Старение листьев и листопад
Листопаду предшествует старение листьев. Это значит, что во всех клетках снижается интенсивность жизненных процессов — фотосинтеза, дыхания. Уменьшается содержание уже имеющихся в клетках важных для растения веществ и сокращается поступление новых, в том числе и воды. Распад веществ преобладает над их образованием. В клетках накапливаются ненужные, и даже вредные продукты, их называют конечными продуктами обмена веществ. Эти вещества удаляются из растения при сбрасывании листьев. Наиболее же ценные соединения по проводящим тканям оттекают из листьев в другие органы растения, где откладываются в клетках запасающих тканей или сразу используется организмом для питания.
У большинства деревьев и кустарников в период старения листья меняют окраску и становятся жёлтыми или багряными. Это происходит потому, что хлорофилл разрушается. Но помимо него в пластидах (хлоропластах) имеются вещества желтого и оранжевого цвета. Летом они были, как бы замаскированы хлорофиллом и пластиды имели зелёный цвет. Кроме того, в вакуолях накапливаются другие красящие вещества жёлтого или красно-малинового цвета. Вместе с пигментами пластид они определяют окраску осенних листьев. У некоторых растений листья сохраняют зелёный цвет до отмирания.
Ещё до того как с побега упадёт лист, в его основании на границе со стеблем формируется слой пробки. Наружу от него образуется отделительный слой. Со временем клетки этого слоя оделяются друг от друга, так как ослизняется и разрушается межклеточное вещество, которое их соединяло, а иногда и оболочки клеток. Лист отделяется от стебля. Однако некоторое время он ещё сохраняется на побеге благодаря проводящим пучкам между листом и стеблем. Но наступает момент нарушения и этой связи. Рубец на месте отделившегося листа покрыт защитной тканью, пробкой.
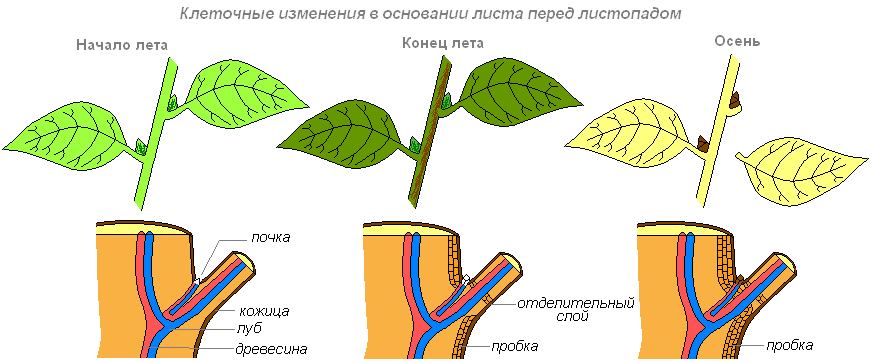
Как только листья достигают предельных размеров, начинаются процессы старения, ведущие, в конце концов, к отмиранию листа — его пожелтение или покраснение, связанное с разрушением хлорофилла, накоплением каротиноидов и антоцианов. По мере старения листа снижается также интенсивность фотосинтеза и дыхания, деградируют хлоропласты, накапливаются некоторые соли (кристаллы оксалаты кальция), из листа оттекают пластические вещества (углеводы, аминокислоты).
В процессе старения листа близ его основания у двудольных древесных растений формируется так называемый отделительный слой, который состоит из легко расслаивающейся паренхимы. По этому слою лист и отделяется от стебля, причём на поверхности будущего листового рубца заранее образуется защитный слой пробковой ткани.

На листовом рубце заметны в виде точек поперечные сечения листового следа. Скульптура листового рубца различна и является характерным признаком для систематики лепидофитов.
У однодольных и травянистых двудольных отделительный слой, как правило, не образуется, лист отмирает и разрушается постепенно, оставаясь на стебле.
У листопадных растений опадение листьев на зиму имеет приспособительное значение: сбрасывая листья, растения резко уменьшают испаряющую поверхность, защищаются от возможных поломок под тяжестью снега. У вечнозелёных растений массовый листопад приурочен обычно к началу роста новых побегов из почек и поэтому происходит не осенью, а весной.
Осенний листопад в лесу имеет важное биологическое значение. Опавшие листья — хорошее органическое и минеральное удобрение. Ежегодно в на их лиственных лесах опавшие листья служат материалом для минерализации, производимой почвенными бактериями и грибами. Кроме того, опавшая листва стратифицирует семена, опавшие до листопада, предохраняет корни от вымерзания, препятствует развитию мохового покрова и т.д. некоторые виды деревьев сбрасывают не только листву, но и годовалые побеги.
Хотя учёные давно знали об испарении воды поверхностью листа, первым, кто наблюдал устьица, был итальянский натуралист Марчелло Мальпиги , который это открытие опубликовал в 1675 году в своей работе Anatome plantarum . Однако он не понял их настоящую функцию. В то же время его современник Неемия Грю развил гипотезу об участии устьиц в вентиляции внутренней среды растения и сравнил их с трахеями насекомых . Прогресс в изучении наступил в XIX веке , и тогда же, в 1827 году , швейцарским ботаником Декандолем было впервые использовано слово „stoma“. Изучением устьиц в то время занимались Гуго фон Моль , который открыл основной принцип открывания устьиц и Симон Швенденер , классифицировавший устьица по типу их конструкции.
Некоторые аспекты функционирования устьиц продолжают интенсивно изучаться и в настоящее время; материалом в основном служат Коммелина обыкновенная (Commelina communis ), Боб садовый (Vicia faba ), Кукуруза сахарная (Zea mays ) .
Строение
Размеры устьица (длина) колеблются в пределах 0,01-0,06 мм (крупнее бывают устьица полиплоидных растений и у листьев, растущих в тени. Самые крупные устьица были обнаружены у вымершего растения Zosterophyllum , 0,12 мм (120 мкм) . Пора состоит из пары специализированных клеток, называемых замыкающими (cellulae claudentes ), которые регулируют степень открытости поры, между ними располагается устьичная щель (porus stomatalis ). Стенки замыкающих клеток утолщены неравномерно: направленные к щели (брюшные) толще стенок, направленных от щели (спинных). Щель может расширяться и сужаться, регулируя транспирацию и газообмен. Когда воды мало, замыкающие клетки плотно прилегают друг к другу и устьичная щель закрыта. Когда воды в замыкающих клетках много, то она давит на стенки и более тонкие стенки растягиваются сильнее, а более толстые втягиваются внутрь, между замыкающими клетками появляется щель . Под щелью расположена подустьичная (воздушная) полость, окружённая клетками мякоти листа, через которую непосредственно и происходит газообмен . Воздух, содержащий диоксид углерода (углекислый газ) и кислород , проникает внутрь ткани листа через эти поры, и далее используется в процессе фотосинтеза и дыхании. Избыточный кислород, произведённый в процессе фотосинтеза внутренними клетками листа, выходит обратно в окружающую среду через эти же поры. Также, в процессе испарения через поры выделяются пары воды. Клетки эпидермиса , примыкающие к замыкающим, получили название сопровождающих (побочных, соседних, околоустьичных). Они участвуют в движении замыкающих клеток. Замыкающие и сопровождающие клетки образуют устьичный комплекс (устьичный аппарат). Наличие или отсутствие устьиц (видимые части устьиц называют устьичными линиями ) часто используют при классификации растений.
Типы устьиц
Число сопровождающих клеток и их расположение относительно устьичной щели позволяют выделить ряд типов устьиц:
- аномоцитный - сопровождающие клетки не отличаются от остальных клеток эпидермиса , тип весьма обычен для всех групп высших растений , за исключением хвойных ;
- диацитный - характеризуется только двумя сопровождающими клетками, общая стенка которых находится под прямым углом к замыкающим клеткам;
- парацитный - сопровождающие клетки располагаются параллельно замыкающим и устьичной щели;
- анизоцитный - замыкающие клетки окружены тремя сопровождающими, одна из которых заметно крупнее или мельче остальных, такой тип обнаружен только у цветковых растений;
- тетрацитный - четыре сопровождающие клетки, характерен для однодольных ;
- энциклоцитный - сопровождающие клетки образуют узкое колесо вокруг замыкающих клеток;
- актиноцитный - несколько сопровождающих клеток, радиально расходящихся от замыкающих клеток;
- перицитный - замыкающие клетки окружены одной побочной сопровождающей клеткой, устьице не соединено с сопровождающей клеткой антиклинальной клеточной стенкой;
- десмоцитный - замыкающие клетки окружены одной сопровождающей клеткой, устьице соединено с ней антиклинальной клеточной стенкой;
- полоцитный - замыкающие клетки окружены одной сопровождающей не полностью: к одному из устьичных полюсов примыкает одна или две эпидермальные клетки; устьице прикреплено к дистальной стороне единственной сопровождающей клетки, имеющей U-образную или подковообразную форму;
- стефаноцитный - устьице, окружённое четырьмя или более (обычно пять-семь) слабодифференцированными сопровождающими клетками, образующими более или менее отчётливую розетку;
- латероцитный - такой тип устьичного аппарата рассматривается большинством ботаников как простая модификация аномоцитного типа.
Расположение устьиц
Двудольные растения, как правило, в нижней части листа имеют больше устьиц, чем в верхней. Это объясняется тем, что верхняя часть горизонтально-расположенного листа, как правило, лучше освещена, и меньшее количество устьиц в ней препятствует избыточному испарению воды. Листья с устьицами, расположенными на нижней стороне, называются гипостоматическими.
У однодольных растений наличие устьиц в верхней и нижней части листа различно. Очень часто листья однодольных растений расположены вертикально, и в этом случае количество устьиц на обоих частях листа может быть одинаково. Такие листья называются амфистоматическими.
У плавающих листьев на нижней части листа устьица отсутствуют, так как они могут впитывать воду через кутикулу . Листья с устьицами, расположенными на верхней стороне, называются эпистоматическими. У подводных листьев устьица отсутствуют совсем.
Устьица хвойных растений обычно спрятаны глубоко под эндодермой, что позволяет сильно снизить расход воды зимой на испарение, а летом - во время засухи.
У мхов (исключение антоцеротовые) настоящие устьица отсутствуют.
Устьица также различаются по уровню расположения относительно поверхности эпидермиса. Некоторые из них расположены вровень с другими эпидермальными клетками, другие подняты выше или погружены ниже поверхности. У однодольных, листья которых растут преимущественно в длину, устьица образуют правильные параллельные ряды, тогда как у двудольных они располагаются беспорядочно.
Углекислый газ
Так как углекислый газ является одним из ключевых реагентов в процессе фотосинтеза, у большинства растений устьица в дневное время открыты. Проблема состоит в том, что при входе воздух смешивается с парами воды, испаряющимися из листа, и поэтому растение не может получить углекислый газ, одновременно не потеряв некоторое количество воды. У многих растений существует защита от испарения воды в виде закупоривающих устьица восковых отложений.
Когда вы изучали строение отдельных органов и тканей, то в основном знакомились с теневыносливыми растениями, живущими в местах с достаточным увлажнением. Это наиболее типичная экологическая группа растений. Понимая общие закономерности строения и жизнедеятельности растений, зная характеристику этой экологической группы, вы сможете легче разобраться в особенностях организации других групп.
Светолюбивые растения . Свет оказывает большое влияние на форму растений. Растущие на открытом месте светолюбивые деревья, как правило, невысокие, разветвленные, с широкой кроной (сосна, береза, лиственница, белая акация). В лесу эти же деревья выглядят совершенно иначе, например сосна здесь имеет высокий, стройный ствол с кроной, расположенной на самой вершине [ 190]. А у сосны, выросшей на опушке, крона однобокая, годичные кольца древесины шире на освещенной стороне. Под влиянием затенения светолюбивые растения теряют боковые ветви.
Благодаря интенсивному освещению некоторые светолюбивые травянистые растения образуют низкорослые, часто розе- точные формы с укороченным стеблем (подорожник, одуванчик, горные растения), другие растения могут иметь достаточно высокий стебель (луговые травы, иван чай).
Светолюбивые растения имеют характерное строение листьев. Они обычно небольшие, плотные, с блестящей толстой кожицей и многочисленными устьицами. У многих растений листья покрыты восковым налетом или волосками, что предохраняет их от прямого воздействия солнечных лучей. У некоторых растений листья на стебле расположены вертикально (степные злаки) или повернуты ребром к падающим лучам солнца (эвкалипт). Это также избавляет лист от чрезмерного нагревания.
В клетках мякоти листа хлоропластов, как правило, немного, поэтому они имеют светло-зеленую окраску.
У светолюбивых растений хорошо развиты механические ткани и корневая система.
Тенелюбивые растения растут под пологом леса, в глубоких расщелинах и других местах, куда не проникают прямые лучи солнца (лишайник, вороний глаз, ветреница). В таких местах обычно повышенная влажность. Эти условия оказывают влияние на строение тенелюбивых растений.
Механические и проводящие ткани развиты слабо, поэтому побеги обычно хрупкие и нежные. Листовые пластинки у этих растений довольно крупные, тонкие. Кожица листа тонкая, ее клетки часто содержат хлоропласты. Устьица могут быть расположены на верхней и на нижней сторонах листа.
Хлоропласты в клетках листьев крупные, хлорофилла в них больше, чем у светолюбивых растений. Это обеспечивает возможность фотосинтеза при слабом, рассеянном освещении.
Растения водных и избыточно увлажненных мест обитания. Большинство живущих в воде растений имеет очень большую поверхность тела по отношению к его общей массе. Они поглощают воду и растворенные в ней вещества всей поверхностью тела, в связи с чем корневая система у них развита слабо, а иногда и совсем отсутствует (элодея, перистолистник). На погруженных в воду частях растений покровные ткани развиты слабо. Устьиц на подводных листьях нет.
У извлеченных из воды растений стебли и листья обвисают. Это связано с тем, что их механические ткани развиты очень слабо и растения могут сохранять вертикальное положение только в воде, которая их поддерживает.
Иное строение у листьев водных растений, плавающих на поверхности . На их верхней стороне, как и у сухопутных растений, образуются многочисленные устьица (на 1 мм 2 верхней стороны листа кувшинки их 460-500), обеспечивающие газообмен с воздушной средой и интенсивное испарение. Поверхность листа покрыта плотной глянцевой кожицей, она плохо смачивается, поэтому вода скатывается и не заливает устьица.
У всех водных растений сильно развита система межклетников, заполненных воздухом, а у некоторых образуется воздухоносная ткань.
Сухопутные растения, живущие в сильно увлажненной среде, совсем не выносят даже кратковременного иссушения. Это объясняется тем, что они плохо регулируют испарение, не могут сохранять в тканях необходимое количество воды и при засухе вянут и погибают. Среди этих растений можно выделить две группы. К первой относят растения, живущие в сильно увлажненных тенистых местах (в лесу это некоторые виды папоротников, кислица). Для растений этой группы характерны признаки тенелюбивых растений. У них обычно плохо развита корневая система, многочисленные устьица расположены на верхней и на нижней сторонах листа. У некоторых, кроме устьиц, есть особые образования - гидатоды (от греческих слов «гидор» - вода и «одос» - путь), через которые излишки воды удаляются из растений. Поэтому у многих растений, живущих при повышенной влажности почвы и воздуха, на листьях часто появляются капли воды. Это явление получило название «плач растений».
Ко второй группе относят растения, живущие в хорошо освещенных сырых местах, на переувлажненных почвах, во влажном воздухе, например калужница болотная, пушица, подмаренник болотный, рогоз.
Эти растения плохо переносят засуху, ветры. Корневая система у них обычно развита слабо, расположена поверхностно. Из-за недостатка воздуха во влажной почве в корнях и стеблях часто образуется система межклетников, развивается воздухоносная ткань. Некоторые особенности строения этих растений позволяют им быть более устойчивыми к действию прямых солнечных лучей. Кожица их листьев достаточно плотная, устьица расположены главным образом на нижней стороне. Механические ткани хорошо развиты.
Растения сухих мест обитания живут при значительном недостатке влаги, чаще всего в степях, полупустынях и пустынях. Растения этой экологической группы обычно обладают хорошо развитой корневой системой, многие из них запасают воду в тканях корня, стебля или листьев . Их листья имеют толстую, плотную кожицу, опушение или превращены в колючки (кактусы), устьиц немного, и расположены они, как у олеандра, - в углублениях . На листьях и других органах есть водонепроницаемый восковой налет (сахарный тростник), листья в период засухи могут быть свернуты в трубку (ковыль)
Таким образом, среди растений можно выделить несколько основных экологических групп: светолюбивые, теневыносливые, тенелюбивые, растения вод них а избыточно увлажненных мест обитания, растения мест достаточного увлажнения и растения сухих мест обитания. У растений каждой группы выработались приспособления, позволяющие им нормально расти и развиваться в определенных условиях обитания.
Клетки эпидермиса почти непроницаемы для воды и газов благодаря своеоб-разному устройству их наружной стенки. Как же осуществляются газообмен между растением и наружной средой и испарение воды — процессы, не-обходимые для нормальной жизнедеятельности растения? Среди клеток эпидермиса встречаются характерные образования, называемые устьи-цами .
Устьице — щелевидное отверстие, окаймленное с двух сторон двумя замыкающими клетками, имеющими большей частью полулун-ную форму.
Клетки эти живые и со-держат хлорофилловые зерна и кру-пинки крахмала, отсутствующие в других клетках эпидермиса. Особен-но много устьиц на листе. Вид усть-ица с поверхности и в разрезе изоб-ражен на рисунке 40. На поперечном разрезе видно, что непосредственно под устьицем внутри ткани листа находится полость, называемая дыхательной . В пределах щели за-мыкающие клетки более сближены в средней части клеток, а выше и ниже они дальше отступают друг от друга, образуя пространства, называемые передним и задним дво-риком .
Замыкающие клетки способны увеличивать и сокращать свои разме-ры, благодаря чему устьичная щель то широко раскрывается, то сужива-ется или даже совсем бывает закрыта.
Таким образом, замыкающие клетки являются аппаратом, регулирующим процесс открывания и закрывания устьиц.
Как же осуществляется этот процесс?
Стенки замыкающих клеток, обращенные к щели, утолщены значи-тельно сильнее, чем стенки, обращенные к соседним клеткам эпидер-миса. Когда растение освещено и имеет избыток влаги, в хлорофилловых зернах замыкающих клеток происходит накопление крахмала, часть ко-торого превращается в сахар. Сахар, растворенный в клеточном соке, при-тягивает воду из соседних клеток эпидермиса, вследствие чего в замы-кающих клетках повышается тургор. Сильное давление приводит к выпячиванию стенок клеток, примыкающих к эпидермальным, а про-тивоположные, сильно утолщенные стенки выпрямляются. Вследствие этого устьичная щель раскрывается, и газообмен, а также испарение воды увеличиваются. В темноте или при недостатке влаги тур-горное давление уменьшается, замыкающие клетки принимают прежнее по-ложение и утолщенные стенки смыкаются. Щель устьица закрывается.
Устьица расположены на всех молодых неодревесневших наземных ор-ганах растения. Особенно много их на листьях, причем здесь они располо-жены, главным образом, на нижней поверхности. Если лист расположен вертикально, то устьица развиваются с обеих его сторон. У плавающих на поверхности воды листьев некоторых водных растений (например, кувшинки, кубышки) устьица расположены только на верхней стороне листа. Материал с сайта
Число устьиц на 1 кв. мм листовой поверхности в среднем равно 300, однако иногда достигает 600 и более. У рогоза (Typha ) насчитывают свыше 1300 устьиц на 1 кв. мм. Листья, погруженные в воду, устьиц не имеют. Расположены устьица чаще всего равномерно по всей поверхности кожи-цы, но у некоторых растений собраны группами. У однодольных растений, а также на хвоинках многих хвойных они расположены продольными рядами. У растений засушливых областей неред-ко устьица бывают погружены в ткань листа.
Развитие устьиц обычно происхо-дит следующим образом. В отдельных клетках эпидермиса образуются дуго-образные стенки, разделяющие клетку на несколько более мелких так, что центральная из них становится родо-начальницей устьиц. Эта клетка раз-деляется продольной (по оси клетки) перегородкой. Затем эта перегородка расщепляется, и образуется щель. Ог-раничивающие ее клетки становятся замыкающими клетками устьица. У не-которых печеночных мхов имеются свое-образные устьица, лишенные замыкаю-щих клеток.
На этой странице материал по темам:
Устьица представляют собой высокоспециализированные образования эпидермы, состоящие из двух замыкающих клеток, между которыми имеется своеобразный межклетник, или устьичная щель ( рис. 37). Щель может расширяться и сужаться, регулируя транспирацию и газообмен. Под щелью располагается дыхательная, или воздушная, полость, окруженная клетками мякоти листа . Клетки эпидермы, примыкающие к замыкающим, получили название побочных, или околоустьичных. Они участвуют в движении замыкающих клеток. Замыкающие и побочные клетки образуют устьичный аппарат. Число побочных клеток и их расположение относительно устьичной щели позволяют выделять ряд устьичных типов. Их изучением занимается стоматография. Данные стоматографии нередко используются в систематике растений для уточнения систематического положения таксонов. Наиболее часто встречающиеся устьичные типы показаны на рисунке 38 .
Аномоцитный тип устьичного аппарата обычен для всех групп высших растений , исключая хвойные . Побочные клетки в этом случае не отличаются от остальных клеток эпидермы. Диацитный тип характеризуется только двумя побочными клетками, общая стенка которых перпендикулярна устьичной щели. Этот тип обнаружен у некоторых цветковых , в частности у большинства губоцветных и гвоздичных . При парацитном типе побочные клетки располагаются параллельно замыкающим и устьичной щели. Он найден у папоротников , хвощей и ряда цветковых растений . Анизоцитный тип обнаружен только у цветковых растений . Здесь замыкающие клетки окружены тремя побочными, одна из которых заметно крупнее или мельче остальных. Тетрацитным типом устьичного аппарата характеризуются преимущественно однодольные . При энциклоцитном типе побочные клетки образуют узкое кольцо вокруг замыкающих клеток. Подобная структура найдена у папоротников , голосеменных и ряда цветковых . Расположение замыкающих клеток относительно прочих клеток эпидермы у разных видов неодинаково. В одних случаях замыкающие клетки находятся на одном уровне с эпидермальными, иногда выступают над ними или, напротив, залегают значительно глубже (погруженные устьица). Последнее наблюдается у растений, приспособленных к засушливым условиям. Иногда углубления, в которых располагаются устьица, выстланы или прикрыты волосками. Называются они устьичными криптами .
Число и распределение устьиц на листе или побеге варьируют в зависимости от вида растений и условий жизни. Число их обычно колеблется от нескольких десятков до нескольких сотен на 1 кв.мм поверхности.
Механизм движения замыкающих клеток весьма сложен и неодинаков у разных видов. У большинства растений при недостаточном водоснабжении в ночные часы, а иногда и днем тургор в замыкающих клетках понижается и щель замыкается, снижая тем самым уровень транспирации. С повышением тургора устьица открываются. Считают, что главная роль в этих изменениях принадлежит ионам калия. Существенное значение в регуляции тургора имеет присутствие в замыкающих клетках хлоропластов . Первичный крахмал хлоропластов , превращаясь в сахар , повышает концентрацию клеточного сока. Это способствует притоку воды из соседних клеток и переходу замыкающих клеток в упругое состояние.
Общая площадь устьичных отверстий составляет лишь 1-2% площади листа. Несмотря на это, транспирация при открытых устьичных щелях достигает 50-70% испарения, равного по площади открытой водной поверхности.
Публикации по теме
-
.jpg) Безопасная защита растений от болезней и вредителей в июле и августе
Безопасная защита растений от болезней и вредителей в июле и августе
Еще наши предки знали, что хороший урожай зависит не только от усердной и ответственной работы, но и фаз Луны. Узнайте и вы благоприятные...
-
 Рекордный урожай зерновых приведет к дефляции Уборка зерновых в рф году
Рекордный урожай зерновых приведет к дефляции Уборка зерновых в рф году
18.07.2017 - 21:03 Новости Беларуси. Массовая уборка зерновых началась на юго-западе страны, сообщили в программе Новости «24 часа» на...