
Коя тъкан съдържа кръвоносни съдове. Проводима тъкан: структурни характеристики. Каква е разликата между ксилемата на различните растения

Значение и разнообразие от проводими тъкани
Проводимите тъкани са основен компонент на повечето висши растения. Те са задължителен структурен компонент на вегетативните и репродуктивните органи на спорите и семенни растения. Проводимите тъкани в комбинация с клетъчните стени и междуклетъчните пространства, някои клетки на основния паренхим и специализирани предавателни клетки образуват проводяща система, която осигурява далечно и радиално транспортиране на вещества. Поради специалния дизайн на клетките и тяхното местоположение в тялото на растенията, проводящата система изпълнява многобройни, но взаимосвързани функции:
1) движението на водата и минералите, абсорбирани от корените от почвата, както и органична материяобразувани в корените, в стъблото, листата, репродуктивните органи;
2) движението на продуктите на фотосинтезата от зелените части на растението до местата на тяхното използване и съхранение: до корените, стъблата, плодовете и семената;
3) движението на фитохормоните в цялото растение, което създава определен техен баланс, който определя скоростта на растеж и развитие на вегетативните и репродуктивните органи на растенията;
4) радиален транспорт на вещества от проводящи тъкани до съседни живи клетки на други тъкани, например до асимилиращи клетки на листния мезофил и делящи се меристемни клетки. В него могат да участват и паренхимните клетки на медуларните лъчи на дървото и кората. Голямо значение в радиалния транспорт имат предавателните клетки с многобройни издатини на клетъчната мембрана, разположени между проводимите и паренхимните тъкани;
5) проводимите тъкани повишават устойчивостта на растителните органи към деформиращи натоварвания;
6) проводимите тъкани образуват непрекъсната разклонена система, която свързва растителните органи в едно цяло;
Появата на проводими тъкани е резултат от еволюционни структурни трансформации, свързани с появата на растенията на сушата и разделянето на тяхното въздушно и почвено хранене. Най-древните проводими тъкани, трахеидите, са открити във фосилни ринофити. Най-високото си развитие са достигнали при съвременните покритосеменни растения.
В процеса на индивидуално развитие се формират първични проводящи тъкани от прокамбий в точките на растеж на семенния ембрион и възобновителните пъпки. Вторичните проводими тъкани, характерни за двусемеделните покритосеменни растения, се генерират от камбия.
В зависимост от изпълняваните функции проводимите тъкани се разделят на тъкани с възходящ ток и тъкани с низходящ ток. Основното предназначение на тъканите на възходящото течение е транспортирането на водата и разтворените в нея минерали от корена към по-високите надземни органи. Освен това те преместват органични вещества, образувани в корена и стъблото, например органични киселини, въглехидрати и фитохормони. Терминът "възходящо течение" обаче не трябва да се приема еднозначно като движение отдолу нагоре. Тъканите с възходящ поток осигуряват поток от вещества в посока от зоната на засмукване към върха на издънката. В този случай транспортираните вещества се използват както от самия корен, така и от стъблото, клоните, листата, репродуктивните органи, независимо дали са над или под нивото на корените. Например, картофите имат вода и елементи минерално храненете навлизат през тъканите на възходящото течение в образуваните в почвата столони и грудки, както и в надземните органи.
Тъканите с низходящ поток осигуряват изтичането на продукти от фотосинтезата към растящите части на растенията и органите за съхранение. В този случай пространственото разположение на фотосинтезиращите органи няма значение. Например при пшеницата органичните вещества навлизат в развиващите се зърна от листа от различни нива. Следователно наименованията "възходящи" и "низходящи" тъкани трябва да се третират като нищо повече от установена традиция.
Проводима нагоре тъкан
Тъканите на възходящия поток включват трахеиди и съдове (трахеи), които се намират в дървесната (ксилемна) част на растителните органи. В тези тъкани движението на водата и разтворените в нея вещества се извършва пасивно под действието на натиска на корена и изпарението на водата от повърхността на растението.
Трахеидите са с по-древен произход. Те се намират във висшите спорови растения, голосеменни и по-рядко при покритосеменни. При покритосеменните те са типични за най-малкото разклонение на листните вени. Трахеидните клетки са мъртви. Те са удължени, често вретеновидни. Дължината им е 1 - 4 мм. Но при голосеменните, като араукарията, тя достига 10 mm. Клетъчните стени са дебели, целулозни, често импрегнирани с лигнин. Клетъчните мембрани имат множество оградени пори.
Съдове, образувани на по-късни етапи от еволюцията. Те са характерни за покритосеменните растения, но се срещат и при някои съвременни представители на отделите Клубни мъхове (род Sellaginella), хвощове, папратовидни и голосеменни (род Gnetum).
Съдовете се състоят от продълговати мъртви клетки, разположени една над друга и се наричат съдови сегменти. В крайните стени на съдовите сегменти има големи проходни отвори - перфорации, през които се осъществява транспортирането на вещества на дълги разстояния. Перфорациите са възникнали в хода на еволюцията от ръбестите пори на трахеидите. Като част от съдовете те са стълбовидни и прости. По крайните стени на съдовите сегменти при наклонено полагане се образуват многобройни стълбовидни перфорации. Отворите на такива перфорации имат удължена форма, а преградите, които ги разделят, са успоредни една на друга, наподобяващи стъпалата на стълба. Съдовете със стълбовидна перфорация са типични за растенията от семействата Ranunculaceae, Schisandra, Birch, Palm, Chastukhove.
Простите перфорации са известни в еволюционно по-млади семейства, като Solanaceae, Cucurbitaceae, Asteraceae и Bluegrass. Те представляват един голям отвор в крайната стена на сегмента, разположен перпендикулярно на оста на съда. В редица семейства, например при Magnoliaceae, Roses, Irises, Asters, в съдовете се срещат както прости, така и стълбовидни перфорации.
Страничните стени имат неравномерни целулозни удебеления, които предпазват съдовете от прекомерно налягане, създадено от съседни живи клетки на други тъкани. В страничните стени може да има множество пори, които осигуряват изхода на водата извън съда.
В зависимост от естеството на удебеленията, вида и естеството на разположението на порите, съдовете се разделят на пръстеновидни, спирални, биспирални, мрежести, стълбовидни и точковидни. В пръстеновидни и спирални съдове целулозните удебеления са подредени под формата на пръстени или спирали. Дифузията на транспортираните разтвори в околните тъкани се осъществява през неудебелените зони. Диаметърът на тези съдове е относително малък. В ретикуларните, скалариформените и точковите пори съдове цялата странична стена, с изключение на местата на прости пори, е удебелена и често импрегнирана с лигнин. Поради това радиалният транспорт на веществата в тях се осъществява през множество удължени и точковидни пори.
Съдовете имат ограничено времедейности. Те могат да бъдат унищожени в резултат на блокиране от тили - израстъци на съседни паренхимни клетки, както и под действието на центростремителни сили на натиск на нови дървесни клетки, образувани от камбия. В хода на еволюцията съдовете претърпяват промени. Съдовите сегменти стават по-къси и по-дебели, наклонените напречни прегради се заменят с прави, а перфорациите на стълбата са прости.
Проводими тъкани надолу
Тъканите надолу по веригата включват ситовидни клетки и ситовидни тръби с придружаващи клетки. Ситовите клетки са с по-древен произход. Срещат се във висшите спорови растения и голосеменните. Това са живи, продълговати клетки със заострени краища. В зряло състояние съдържат ядра в състава на протопласта. В страничните им стени, в местата на контакт на съседни клетки, има малки проходни отвори, които са събрани в групи и образуват ситовидни полета, през които се осъществява движението на веществата.
Ситовите тръби се състоят от вертикален ред удължени клетки, разделени от напречни стени и наречени ситови плочи, в които са разположени ситови полета. Ако ситовата плоча има едно ситово поле, то се счита за просто, а ако има няколко, тогава е сложно. Ситовите полета са оформени от множество проходни отвори - перфорации на сито с малък диаметър. Плазмодесматите преминават през тези перфорации от една клетка в друга. По стените на перфорациите се поставя калозен полизахарид, който намалява лумена на перфорациите. Тъй като ситовата тръба старее, калозата напълно запушва перфорациите и тръбата спира да работи.
Когато се образува ситовидна тръба, в клетките, които ги образуват, се синтезира специален флоемен протеин (Р-протеин) и се развива голяма вакуола. Той избутва цитоплазмата и ядрото към клетъчната стена. След това вакуолната мембрана се разпада и вътрешно пространствоКлетките са пълни със смес от цитоплазма и клетъчен сок. F-протеиновите тела губят отчетливите си очертания, сливат се, образувайки нишки близо до ситовите плочи. Техните фибрили преминават през перфорации от един сегмент на ситовата тръба в друг. Една или две сателитни клетки, които имат удължена форма, тънки стени и жива цитоплазма с ядро и множество митохондрии, са плътно прикрепени към сегментите на ситовидната тръба. АТФ се синтезира в митохондриите, което е необходимо за транспортирането на вещества през ситовидните тръби. В стените на сателитните клетки има голям брой пори с плазмадесми, който е почти 10 пъти по-голям от техния брой в други клетки на мезофила на листата. Повърхността на протопласта на тези клетки е значително увеличена поради множество гънки, образувани от плазмалемата.
Скоростта на движение на асимилатите през ситовите тръби значително надвишава скоростта на свободната дифузия на веществата и достига 50–150 cm / h, което показва активен транспорт на вещества с помощта на енергията на АТФ.
Продължителността на ситовидните тръби при многогодишните двусемеделни растения е 1 - 2 години. За да ги замени, камбият постоянно образува нови проводими елементи. При едносемеделните без камбий ситовидните тръби съществуват много по-дълго.
Проводими снопове
Проводимите тъкани са разположени в растителните органи под формата на надлъжни нишки, образуващи съдови снопове. Има четири вида съдови снопове: прости, общи, сложни и съдово-фиброзни.
Простите снопове се състоят от един вид проводяща тъкан. Например, в крайните части на листните остриета на много растения се откриват снопове от съдове и трахеиди с малък диаметър, а в цъфтящите издънки на лилии се откриват само ситови тръби.
Общите снопове се образуват от трахеиди, съдове и ситовидни тръби. Понякога терминът се използва за обозначаване на снопчета метамери, които се движат в междините и са следи от листа. Сложните снопове включват проводими и паренхимни тъкани. Най-съвършените, разнообразни по структура и местоположение са съдово-фиброзните снопове.
Съдовите влакнести снопове са характерни за много висши спорови растения и голосеменни. Те обаче са най-характерни за покритосеменните. В такива снопове се разграничават функционално различни части - флоема и ксилема. Флоемата осигурява изтичането на асимилатите от листата и придвижването им до местата на използване или съхранение. Водата и разтворените в нея вещества се движат по ксилемата от кореновата система към листата и други органи. Обемът на ксилемната част е няколко пъти по-голям от обема на флоемната част, тъй като обемът на водата, постъпваща в растението, надвишава обема на образуваните асимилати, тъй като значителна част от водата се изпарява от растението.
Разнообразието от съдови влакнести снопове се определя от техния произход, хистологичен състав и местоположение в растението. Ако сноповете се образуват от прокамбиума и завършат развитието си, тъй като доставката на клетки от образователната тъкан се изразходва, както при едносемеделните, те се наричат затворени за растеж. За разлика от това, при двусемеделните отворените туфи не са ограничени в растежа, тъй като се образуват от камбия и увеличават диаметъра си през целия живот на растението. Съставът на съдово-фиброзните снопове, в допълнение към проводимите, може да включва основни и механични тъкани. Например при двусемеделните флоемата се образува от ситовидни тръби (проводяща тъкан нагоре), ликов паренхим (основна тъкан) и ликови влакна (механична тъкан). Съставът на ксилемата включва съдове и трахеиди (проводяща тъкан на низходящ поток), дървесен паренхим (основна тъкан) и дървесни влакна (механична тъкан). Хистологичният състав на ксилема и флоема е генетично определен и може да се използва в таксономията на растенията за диагностициране на различни таксони. В допълнение, степента на развитие на съставните части на сноповете може да се промени под влияние на условията на растеж на растенията.
Известни са няколко вида съдово-фиброзни снопове.
Затворените колатерални съдови снопове са характерни за листата и стъблата на едносемеделните покритосеменни растения. Липсва им камбий. Флоема и ксилемата са разположени една до друга. Те се характеризират с някои дизайнерски характеристики. И така, при пшеницата, която се различава по начина на фотосинтеза С 3, сноповете се образуват от прокамбий и имат първичен флоем и първичен ксилем. Във флоема се разграничават по-ранен протофлоем и по-късен по време на образуване, но по-едроклетъчен метафлоем. Във флоемната част липсва личен паренхим и ликови влакна. В ксилемата първоначално се образуват по-малки протоксилемни съдове, разположени в една линия, перпендикулярна на вътрешната граница на флоема. Метаксилемата е представена от два големи съда, разположени до метафлоема перпендикулярно на веригата от протоксилемни съдове. В този случай съдовете са разположени Т-образно. Известна е и V-, Y- и È-образната подредба на съдовете. Между съдовете на метаксилемата в 1-2 реда има дребноклетъчен склеренхим с удебелени стени, които се импрегнират с лигнин, когато се развива стъблото. Този склеренхим разделя зоната на ксилемата от флоема. От двете страни на съдовете на протоксилема има клетки от дървесния паренхим, които вероятно играят трансфузионна роля, тъй като когато пакетът преминава от междувъзлията към листната възглавница на стволовия възел, те участват в образуването на предавателни клетки . Около съдовия сноп на стъблото на пшеницата има склеренхимна обвивка, по-добре развита от страната на протоксилема и протофлоема, близо до страничните страни на снопа, клетките на обвивката са подредени в един ред.
При растения с тип фотосинтеза С 4 (царевица, просо и др.) В листата около затворените съдови снопове е разположена обвивка от големи хлоренхимни клетки.

Отворените странични снопове са характерни за стъблата на двусемеделните растения. Наличието на камбиев слой между флоема и ксилемата, както и отсъствието на склеренхимна обвивка около снопчетата, осигурява тяхното дългосрочно нарастване на дебелина. В частите на ксилема и флоема на такива снопове има клетки от основната и механичната тъкан.
Отворените пакети от обезпечения могат да се формират по два начина. Първо, това са снопове, образувани предимно от прокамбий. След това в тях се развива камбий от клетките на основния паренхим, произвеждайки вторични елементи на флоема и ксилема. В резултат на това сноповете ще комбинират хистологични елементи от първичен и вторичен произход. Такива снопове са характерни за много тревисти цъфтящи растения от класа на двусемеделните, които имат снопов тип структура на стъблото (бобови, розоцветни и др.).
Второ, отворените съпътстващи снопове могат да се образуват само от камбия и се състоят от ксилема и флоема от вторичен произход. Характерни са за тревисти двусемеделни растения с преходен тип анатомичен строеж на стъблото (Asteraceae и др.), както и за кореноплодни култури като цвекло.
В стъблата на растения от редица семейства (Pucuraceae, Solanaceae, Campanaceae и др.) Има отворени двустранни снопове, където ксилемата е заобиколена от двете страни от флоема. В същото време външната част на флоема, обърната към повърхността на стъблото, е по-добре развита от вътрешната, а лентата на камбия, като правило, се намира между ксилемата и външната част на флоема.
Концентричните снопове са два вида. В амфикрибралните снопове, характерни за коренищата на папрата, флоемата обгражда ксилемата, в амфивазалните снопчета ксилемата е разположена в пръстен около флоема (коренищата на ириса, момина сълза и др.). По-рядко концентрични снопове се срещат при двусемеделните растения (рицин).
В участъците на корените, които имат първична анатомична структура, се образуват затворени радиални съдови снопове. Радиалният сноп е част от централния цилиндър и минава през средата на корена. Ксилема му има вид на многолъчева звезда. Флоемните клетки са разположени между лъчите на ксилема. Броят на ксилемните лъчи до голяма степен зависи от генетичната природа на растенията. Например при моркови, цвекло, зеле и други двусемеделни растения ксилемата на радиалния сноп има само две рамена. Ябълката и крушата могат да имат 3-5 от тях, тиквата и бобът имат четирилъчева ксилема, а едносемеделното растение има многолъчева ксилема. Радиалното разположение на ксилемните лъчи има адаптивна стойност. Скъсява пътя на водата от смукателната повърхност на корена до съдовете на централния цилиндър.
Трайни насаждения дървесни растенияи някои тревисти едногодишни, като лен, проводимите тъкани са разположени в стъблото, без да образуват ясно изразени проводящи снопове. Тогава те говорят за нелъчевия тип структура на стъблото.
Тъкани, които регулират радиалния транспорт на веществата
Специфични тъкани, които регулират радиалния транспорт на вещества, включват екзодерма и ендодерма.

Екзодермата е външният слой на първичната коренова кора. Образува се директно под първичната епиблема на покривната тъкан в зоната на кореновите косми и се състои от един или повече слоя плътно затворени клетки с удебелени целулозни мембрани. В екзодермата водата, която е навлязла в корена по кореновите власинки, изпитва съпротивление от вискозната цитоплазма и се премества в целулозните мембрани на клетките на екзодермата и след това ги напуска в междуклетъчните пространства на средния слой на първичната кора, или мезодерма. Това осигурява ефективен поток на вода в по-дълбоките слоеве на корена.
В зоната на проводимост в корена на едносемеделните, където клетките на епиблемата умират и се десквамират, екзодермата се появява на повърхността на корена. Неговите клетъчни стени са импрегнирани със суберин и предотвратяват притока на вода от почвата към корена. При двусемеделните екзодермата в първичната кора се отделя по време на линеене на корените и се заменя с перидерма.
Ендодермата или вътрешният слой на кортекса на първичния корен е разположен около централния цилиндър. Образува се от един слой плътно затворени клетки с неравномерна структура. Някои от тях, наречени пропуски, имат тънки черупкии лесно пропускливи за вода. През тях водата от първичната кора постъпва в радиалния проводящ сноп на корена. Други клетки имат специфични целулозни удебеления на радиалните и вътрешните тангенциални стени. Тези удебеления, импрегнирани със суберин, се наричат каспарови ленти. Те са водонепропускливи. Следователно водата навлиза в централния цилиндър само през проходните клетки. И тъй като абсорбиращата повърхност на корена значително надвишава общата площ на напречното сечение на клетките на пропускливата ендодерма, тогава възниква коренов натиск, който е един от механизмите за навлизане на вода в стъблото, листата и репродуктивните органи.
Ендодермата също е част от кората на младото стъбло. При някои тревисти покритосеменни тя, както и в корена, може да има каспари ленти. В допълнение, при младите стъбла ендодермата може да бъде представена от нишестена обвивка. По този начин ендодермата може да регулира водния транспорт в растението и да съхранява хранителни вещества.
Концепцията за стелата и нейната еволюция
Много внимание се обръща на възникването, развитието в онтогенезата и еволюционните структурни трансформации на проводящата система, тъй като тя осигурява взаимовръзката на растителните органи и еволюцията на големи таксони е свързана с нея.
По предложение на френските ботаници F. Van Tiegem и A. Dulio (1886) съвкупността от първични проводящи тъкани, заедно с други тъкани, разположени между тях и перицикла, съседен на кората, се нарича стела. Съставът на стелата може също да включва ядро и кухина, образувана на негово място, както например в синята трева. Понятието "стела" съответства на понятието "централен цилиндър". Стелата на корена и стъблото е функционално еднаква. Изследването на стелата в представители на различни отдели на висшите растения доведе до формирането на теорията за стелата.
Има два основни вида стела: протостела и еустела. Най-древна е протостелата. Проводимите му тъкани са разположени в средата на аксиалните органи, а в центъра е ксилема, заобиколена от непрекъснат слой флоема. В стъблото няма сърцевина или кухина.
Има няколко еволюционно свързани типа протостела: хаплостела, актиностела и плектостела.
Първоначалният, примитивен вид е хаплостелата. Ксилема й има заоблена форма на напречно сечение и е заобиколена от равномерен непрекъснат слой флоема. Перицикълът е разположен около проводимите тъкани в един или два слоя. Хаплостелата е била известна при изкопаеми ринофити и е запазена при някои псилотофити (tmesipterus).
По-развит тип протостела е актиностелата, в която ксилемата в напречно сечение има формата на многолъчева звезда. Намерено е във вкаменелостите Asteroxylon и някои примитивни ликопсиди.
По-нататъшното разделяне на ксилемата на отделни участъци, разположени радиално или успоредно един на друг, доведе до образуването на плектостела, характерна за стъблата на Lycopod. При актиностела и плектостела флоемата все още обгражда ксилемата от всички страни.
В хода на еволюцията от протостелата възниква сифоностела, чиято отличителна черта е тръбната структура. В центъра на такава стела има ядро или кухина. В проводящата част на сифоностела се появяват листни празнини, поради което има непрекъсната връзка между сърцевината и кората. В зависимост от начина на взаимно разположение на ксилемата и флоема, сифоностела може да бъде ектофлоен и амфифлоичен. В първия случай флоемата обгражда ксилемата от едната, външната страна. Във втория флоемата обгражда ксилемата от две страни, от външната и вътрешната.
Когато амфифлоичната сифоностела е разделена на мрежа или редици от надлъжни нишки, се образува разчленена стела или диктиостела, която е характерна за много папрати. Проводимата му част е представена от множество концентрични проводящи снопове.
При конските опашки артростела възниква от ектофлоичния сифоностел, който има съчленена структура. Отличава се с наличието на една голяма централна кухина и изолирани съдови снопове с протоксилемни кухини (каринални канали).
При цъфтящите растения на базата на ектофлоичната сифоностела се образуват еустела, характерна за двусемеделните, и атактостела, характерна за едносемеделните. В еустелата проводящата част се състои от отделни странични снопове, имащи кръгово разположение. В центъра на стелата в стъблото е сърцевината, която е свързана с кората с помощта на сърцевини. В атактостелата съдовите снопове са разпръснати, като между тях са разположени паренхимни клетки на централния цилиндър. Тази подредба на сноповете скрива тръбната структура на сифонната стела.
възникване различни опции siphonostele е важна адаптация на висшите растения към увеличаване на диаметъра на аксиалните органи - корена и стъблото.
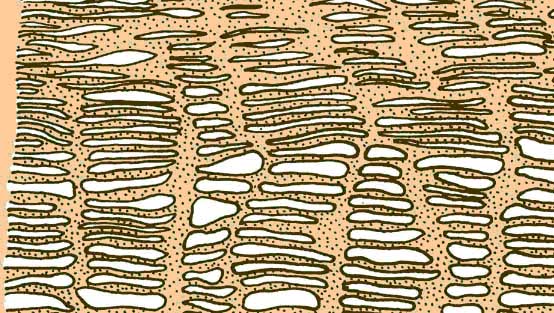
Проводимите тъкани са сложни, тъй като се състоят от няколко вида клетки, техните структури имат удължена (тръбна) форма и са проникнати от множество пори. Наличието на отвори в крайните (долни или горни) секции осигурява вертикален транспорт, а порите на страничните повърхности допринасят за потока на водата в радиална посока. Проводимите тъкани включват ксилема и флоема. Те се срещат само в папрати и семенни растения. Проводимата тъкан съдържа както мъртви, така и живи клетки.
Ксилем (дърво)е мъртва тъкан. Включва основните структурни компоненти (трахеи и трахеиди), дървесен паренхим и дървесни влакна. Той изпълнява както поддържаща, така и проводяща функция в растението - водата и минералните соли се движат нагоре по растението по него.
трахеиди
- мъртви единични клетки с вретеновидна форма. Стените са силно удебелени поради отлагането на лигнин. Характеристика на трахеидите е наличието на оградени пори в стените им. Краищата им се припокриват, давайки на растението необходимата здравина. Водата се движи през празните празнини на трахеидите, без да среща препятствия под формата на клетъчно съдържание по пътя си; от една трахеида на друга, тя се предава през порите.
При покритосеменните трахеидите са се развили в кръвоносни съдове (трахея). Това са много дълги тръби, образувани в резултат на „докинг“ на редица клетки; останките от крайните прегради все още са запазени в съдовете под формата на перфорирани ръбове. Съдовете варират по размер от няколко сантиметра до няколко метра. В първите съдове на образуване на протоксилема лигнинът се натрупва в пръстени или в спирала. Това позволява на съда да продължи да се разтяга по време на растежа. В съдовете на метаксилема лигнинът е концентриран по-плътно - това е идеален "водопровод", който действа на големи разстояния.
?1.
Как се различават трахеите от трахеидите? (Отговор в края на статията)
?2
. Как трахеидите се различават от влакната?
?3
. Какво е общото между флоема и ксилема?
?4.
Как се различават ситовидните тръби от трахеите?
Паренхимните ксилемни клетки образуват своеобразни лъчи, свързващи ядрото с кората. Те провеждат вода в радиална посока, съхраняват хранителни вещества. Нови ксилемни съдове се развиват от други паренхимни клетки. И накрая, дървесните влакна са подобни на трахеидите, но за разлика от тях имат много малък вътрешен лумен, следователно не провеждат вода, но придават допълнителна здравина. Освен това имат прости пори, а не оградени.
Флоема (лико)- това е жива тъкан, който е част от кората на растенията, през него се осъществява низходящ поток от вода с разтворени в нея асимилационни продукти. Флоемата се формира от пет вида структури: ситовидни тръби, придружаващи клетки, личен паренхим, ликови влакна и склереиди.
Тези структури се основават на ситови тръби
, образуван в резултат на свързването на редица ситови клетки. Стените им са тънки, целулозни, ядрата умират след узряване, а цитоплазмата се притиска към стените, освобождавайки място за органични вещества. Крайните стени на клетките на ситовите тръби постепенно се покриват с пори и започват да приличат на сито - това са ситови плочи. За да се осигури тяхната жизнена активност, сателитните клетки са разположени наблизо, тяхната цитоплазма е активна, ядрата са големи.
?5
. Защо мислите, че когато ситовидните клетки узреят, техните ядра умират?
ОТГОВОРИ
?1.
Трахеите имат многоклетъчни структури и нямат крайни стени, докато трахеидите са едноклетъчни, имат крайни стени и оградени пори.
?2
. Трахеидите имат оградени пори и добре дефиниран лумен, докато при влакната луменът е много малък и порите са прости. Те също се различават по функции, трахеидите изпълняват транспортна роля (проводяща) и механични влакна.
?3.
Флоемата и ксилемата са проводими тъкани, техните структури са с тръбна форма, включват клетки на паренхима и механични тъкани.
?4.
Ситовидните тръби се състоят от живи клетки, стените им са целулозни, извършват низходящ транспорт на органични вещества, а трахеите са образувани от мъртви клетки, стените им са силно удебелени с лигнин, осигуряват възходящ транспорт на вода и минерали.
?5.
Низходящият транспорт се извършва по ситовите клетки и ядрата, отнесени от потока на веществата, биха покрили значителна част от ситовото поле, което би довело до намаляване на ефективността на процеса.
Проводимата тъкан се състои от живи или мъртви продълговати клетки, които приличат на тръби.
В стъблото и листата на растенията има снопове проводяща тъкан. В проводящата тъкан се изолират съдове и ситовидни тръби.
Съдове- последователно свързани мъртви кухи клетки, напречните прегради между които изчезват. Чрез съдовете водата и разтворените в нея минерали от корените навлизат в стъблото и листата.
ситови тръби - удължени безядрени живи клетки, свързани последователно една с друга. Чрез тях органичните вещества от листата (където са се образували) се придвижват до други органи на растението.
Проводимата тъкан осигурява транспортиране на вода с разтворени в нея минерали.
Тази тъкан образува две транспортни системи:
- възходящ(от корените до листата);
- низходящ(от листата до всички други части на растенията).
Възходящата транспортна система се състои от трахеиди и съдове (ксилем или дърво), а съдовете са по-съвършени проводящи средства от трахеидите.
В низходящите системи потокът вода с продукти на фотосинтезата преминава през ситови тръби (флоем или лико).
Ксилема и флоемата образуват съдови влакнести снопове - " кръвоносна система„на растение, което го прониква изцяло, свързвайки го в едно цяло.
Учените смятат, че появата на тъканите е свързана в историята на Земята с освобождаването на растенията на сушата. Когато част от растението е в въздушна среда, а другата част (корен) - в почвата, стана необходимо да се доставя вода и минерални соли от корените до листата, а органичните вещества - от листата до корените. Така в хода на еволюцията флоравъзникват два вида проводими тъкани - дърво и лико.
През дървесината (чрез трахеиди и съдове) водата с разтворени минерали се издига от корените към листата - това е водопроводно или възходящо течение. Чрез лика (чрез ситовидни тръби) органичните вещества, образувани в зелените листа, навлизат в корените и другите органи на растението - това е низходящ поток.
образователна тъкан
Образователната тъкан се намира във всички растящи части на растението. Образователната тъкан се състои от клетки, които могат да се делят през целия живот на растението. Клетките тук лежат много бързо една към друга. Благодарение на деленето те образуват много нови клетки, като по този начин осигуряват растежа на растението по дължина и дебелина. Клетките, които се появяват по време на разделянето на образователни тъкани, след това се трансформират в клетки от други растителни тъкани.

Това е първичната тъкан, от която се образуват всички останали растителни тъкани. Състои се от специални клетки, способни на многократно делене. Именно от тези клетки се състои ембрионът на всяко растение.
Тази тъкан се запазва във възрастно растение. Намира се:
- в долната част на кореновата система и по върховете на стъблата (осигурява растежа на растението във височина и развитието на кореновата система) - апикална образователна тъкан;
- вътре в стъблото (осигурява растежа на растението в ширина, неговото удебеляване) - странична образователна тъкан.
За разлика от други тъкани, цитоплазмата на образователната тъкан е по-дебела и по-плътна. Клетката има добре развити органели, които осигуряват синтеза на протеини. Ядрото е голямо. Масата на ядрото и цитоплазмата се поддържат в постоянно съотношение. Увеличаването на ядрото сигнализира за началото на процеса на клетъчно делене, което се осъществява чрез митоза за вегетативните части на растенията и мейоза за спорогенните меристеми.
ПРОВОДЯЩИ ТЪКАНИ
Проводимите тъкани транспортират хранителни вещества в две посоки. Възходящ (транспирационен) токтечност ( водни разтвории сол) върви заедно съдовеи трахеидиксилеми (фиг. 32) от корените нагоре по стъблото до листата и другите органи на растението. Низходящо течение (асимилаторно)органичната материя се пренася от листата по стъблото до подземните органи на растението

специален ситови тръбифлоема (фиг. 33). Проводимата тъкан на растението донякъде напомня на човешката кръвоносна система, тъй като има аксиална и радиална силно разклонена мрежа; Хранителните вещества влизат във всяка клетка на живо растение. Във всеки растителен орган ксилемата и флоемата са разположени един до друг и са представени под формата на нишки - проводящи снопове.
Има първични и вторични проводими тъкани. Първичните се диференцират от прокамбия и се полагат в младите органи на растението, вторичните проводящи тъкани са по-мощни и се образуват от камбия.
Ксилем (дърво)представени трахеидии трахеи, или съдове.
трахеиди- удължени затворени клетки с наклонени назъбени краища, в зряло състояние са представени от мъртви прозенхимни клетки. Дължината на клетките е средно 1 - 4 mm. Комуникацията със съседните трахеиди се осъществява чрез прости или ресни пори. Стените са неравномерно удебелени, по естеството на удебеляването на стените трахеидите са пръстеновидни, спирални, стъпаловидни, мрежовидни и порести (фиг. 34). Порестите трахеиди винаги имат оградени пори (фиг. 35). Спорофитите на всички висши растения имат трахеиди, а в повечето хвощове, ликопсиди, папрати и голосеменни те служат като единствените проводящи елементи на ксилемата. трахеиди


изпълняват две основни функции: провеждане на вода и механично укрепване на тялото.
Трахеята, или съдове, - основните водопроводими елементи на ксилемата покритосеменни растения. Трахеите са кухи тръби, състоящи се от отделни сегменти; има дупки в преградите между сегментите - перфорации, през които се осъществява потокът от течност. Трахеите, подобно на трахеидите, са затворена система: краищата на всяка трахея имат скосени напречни стени с оградени пори. Сегментите на трахеята са по-големи от трахеидите: в диаметър са различни видоверастения от 0,1 - 0,15 до 0,3 - 0,7 мм. Дължината на трахеята е от няколко метра до няколко десетки метра (за лиани). Трахеята се състои от мъртви клетки, но начални етапиобразувания те са живи. Смята се, че трахеята в процеса на еволюция е възникнала от трахеидите.
Съдовете и трахеидите, в допълнение към първичната мембрана, имат предимно вторични удебеления под формата на пръстени, спирали, стълби и др. По вътрешната стена на съдовете се образуват вторични удебеления (виж фиг. 34). Така че в съд с пръстени вътрешните удебеления на стените са под формата на пръстени, разположени на разстояние един от друг. Пръстените са разположени напречно на съда и леко косо. В спирален съд вторичната мембрана е наслоена от вътрешността на клетката под формата на спирала; в мрежест съд неудебелените места на черупката изглеждат като процепи, наподобяващи мрежести клетки; в съда със стълба удебелени места се редуват с неудебелени, образувайки подобие на стълба.
Трахеидите и съдовете - елементи на трахеята - са разпределени в ксилемата по различни начини: на напречно сечение, в плътни пръстени, образуващи пръстеновидно дърво, или разпръснати повече или по-малко равномерно в цялата ксилема, образувайки разпръснато съдово дърво. Вторичната обвивка обикновено е импрегнирана с лигнин, което придава на растението допълнителна здравина, но в същото време ограничава растежа му на дължина.
В допълнение към съдовете и трахеидите, ксилема включва греди елементи, състоящ се от клетки, които образуват сърцевини. Медуларните лъчи се състоят от тънкостенни живи паренхимни клетки, през които хранителните вещества преминават в хоризонтална посока. Ксилема съдържа и живи клетки от дървесния паренхим, които функционират като транспорт на къси разстояния и служат като място за съхранение на резервни вещества. Всички елементи на ксилемата идват от камбия.
Флоема- проводима тъкан, през която се транспортират глюкоза и други органични вещества - продукти на фотосинтезата от листата до местата на тяхното използване и отлагане (до растежни конуси, грудки, луковици, коренища, корени, плодове, семена и др.). Флоемата също може да бъде първична и вторична.

Първичната флоема се образува от прокамбия, вторичната (ликова) от камбия. В първичната флоема няма сърцевини и по-малко мощна система от ситови елементи, отколкото в трахеидите. В процеса на образуване на ситовидната тръба в протопласта на клетките - сегменти на ситовидната тръба се появяват слузни тела, които участват в образуването на слузната връв около ситовидните пластини (фиг. 36). Това завършва формирането на сегмента на ситовата тръба. Функциониращи ситовидни тръби в повечето тревисти растенияедин вегетационен период и до 3-4 години при дървета и храсти. Ситовите тръби се състоят от поредица от удължени клетки, които комуникират една с друга чрез перфорирани прегради - цедка. Обвивките на функциониращите ситовидни тръби не лигнифицират и остават живи. Старите клетки се запушват с така нареченото corpus callosum, след което умират и под натиска на по-младите функциониращи клетки се сплескват.
принадлежи към флоема ликов паренхим, състоящ се от тънкостенни клетки, в които се отлагат резервни хранителни вещества. от сърцевинни лъчивторичен флоем, транспортирането на органични вещества на къси разстояния хранителни вещества- продукти на фотосинтезата.
Проводими снопове- нишки, образувани, като правило, от ксилема и флоема. Ако нишките са в съседство с проводящите снопове

механична тъкан (обикновено склеренхим), тогава такива снопове се наричат съдово фиброзен. В съдовите снопове могат да бъдат включени и други тъкани - жив паренхим, лактифери и др. Съдовите снопчета могат да бъдат пълни, когато има и ксилема, и флоема, и непълни, състоящи се само от ксилема (ксилем, или дървесен, съдов сноп) или флоема (флоема или лико, проводящ сноп).
Проводими снопове, първоначално образувани от прокамбий. Има няколко вида проводими лъчи (фиг. 37). Част от прокамбия може да се запази и след това да се превърне в камбий, тогава снопът е способен на вторично удебеляване. то отворенснопове (фиг. 38). Такива съдови снопове преобладават при повечето двусемеделни и голосеменни. Растенията с отворени гроздове могат да растат на дебелина поради активността на камбия, а дървесните участъци (фиг. 39, 5) са приблизително три пъти по-големи от ликовите участъци (фиг. 39, 2) . Ако по време на диференциацията на проводящия сноп от прокамбиалната връв цялата образователна тъкан се изразходва напълно за образуването на постоянни тъкани, тогава снопът се нарича затворен(фиг. 40). Затворено



съдови снопове се намират в стъблата на едносемеделните. Дървесината и ликът в снопове може да са различни взаимно споразумение. В тази връзка се разграничават няколко вида проводящи снопове: колатерални, двустранни (фиг. 41), концентрични и радиални. Обезпечение, или страничен, - снопове, в които ксилемата и флоемата са съседни една на друга. Двустранно, или двустранен, - снопове, в които две нишки от флоема граничат една до друга с ксилемата. AT концентриченв снопове ксилемната тъкан напълно обгражда флоемната тъкан или обратното (фиг. 42). В първия случай такъв лъч се нарича центрофлоем. Снопове центрофлоема се намират в стъблата и коренищата на някои двусемеделни и едносемеделни растения (бегония, киселец, ирис, много острица и лилии). Папратите ги имат. Също така има



междинни проводящи снопове между затворени колатерали и центрофлоема. намерени в корените радиалнаснопове, в които централната част и лъчите по радиусите са оставени от дърво, а всяка греда от дърво се състои от централни по-големи съдове, постепенно намаляващи по радиус (фиг. 43). Броят на лъчите различни растениянеравномерно. Между дървените греди са разположени лугови зони. Видовете проводящи лъчи са показани схематично на фиг. 37. Проводимите снопове се простират по цялото растение под формата на нишки, които започват в корените и преминават по цялото растение по протежение на стъблото до листата и други органи. В листата те се наричат вени. Главна функциятехните - извършване на низходящи и възходящи течения на вода и хранителни вещества.
В процеса на еволюция, с появата на висшите растения на сушата, те развиват тъкани, които достигат най-голямата си специализация в цъфтящите растения. В тази статия ще разгледаме по-подробно какво представляват растителните тъкани, какви видове съществуват, какви функции изпълняват, както и структурните характеристики на растителните тъкани.
плат наричат групи от клетки, които са подобни по структура и изпълняват еднакви функции.
Основните тъкани на растенията са показани на фигурата по-долу:
Видове, функции и структура на растителните тъкани.


Покривна тъкан на растенията.


Покривна тъкан на растенията - кора
Проводима тъкан на растенията.
| Име на тъканта | Структура | Местоположение | Функции |
| 1. Съдове от дърво - ксилема | Кухи тръби с дървесни стени и мъртво съдържание | Дървесина (ксилема), минаваща по корена, стъблото, листните вени | Провеждане на вода и минерали от почвата към корена, стъблото, листата, цветята |
|
2. Ситовидни тръби от лико - флоема Придружаващи клетки или придружаващи клетки |
Вертикален ред от живи клетки с ситовидни напречни прегради Сестрински клетки на ситови елементи, които са запазили структурата си |
Баст (флоем), разположен по корена, стъблото, листните вени Винаги разположени по дължината на ситовите елементи (придружават ги) |
Пренасяне на органична материя от листа до стъбло, корен, цветя приемам Активно участиепри транспортирането на органични вещества през ситовидните тръби на флоема |
| 3. Проводими съдови снопове | Комплекс от дърво и лик под формата на отделни нишки в билки и непрекъснат масив в дървета | Централен цилиндър на корен и стъбло; вени от листа и цветя | Пренасяне на вода и минерали през дърво; върху лика - органични вещества; укрепване на органите, свързването им в едно цяло |



механична тъкан на растенията.


Свързани публикации
-
.jpg) Безопасна защита на растенията от болести и неприятели през юли и август
Безопасна защита на растенията от болести и неприятели през юли и август
Още нашите предци са знаели, че добрата реколта зависи не само от упорита и отговорна работа, но и от фазите на луната. Разберете и сте благосклонни ...
-
 Рекордна зърнена реколта ще доведе до дефлация Зърнена реколта в Руската федерация
Рекордна зърнена реколта ще доведе до дефлация Зърнена реколта в Руската федерация
18.07.2017 - 21:03 Беларус новини. Започна масова жътва на зърнени култури в югозападната част на страната, съобщиха от "24 часа" на...